



















В нейронных цепях, являющихся хранилищем нашей психики и связывающих отдельные части организма в согласованно функционирующее целое, передача сигналов осуществляется двумя очень разными путями:
- электромагнитный импульс проходит через тело нейрона, начинаясь в месте приема сигнала и заканчиваясь в месте его передачи. О том, как этот процесс происходит, можно прочитать в главе 25 (довольно общие сведения) и в главе 26 (более подробное описание).
- нервные клетки взаимодействуют друг с другом и с их мишенями: мышечными и секреторными клетками, передавая сигнал с помощью молекул особых веществ: нейромедиаторов (или просто медиаторов). Место взаимодействия нейронов называется синапсом. Это совсем другой способ связи, и в этой главе он будет рассмотрен всего лишь на базовом уровне, достаточном для формирования общего понимания процесса.
Именно благодаря синапсам нейроны соединяются в нервные цепи, которые достигают невероятной сложности в головном мозге человека. Сам по себе каждый из 100 миллиардов нейронов, несмотря на их несомненную сложность организации и совершенство функций, способен только на самые простые действия. Вся сложность головного мозга, недоступная для постижения и им самим, и современными компьютерами, основана именно на взаимодействии нейронов, каждый из которых в среднем образует тысячу синапсов.
Нейромедиаторы эволюционно консервативны, то есть остаются неизменными на протяжении миллионов лет эволюции. Некоторые из них присутствовали в клетках наших древних предков и могут быть обнаружены сегодня у самых далеких и эволюционно примитивных родственников — одноклеточных организмов, таких как бактерии и дрожжи, и простых многоклеточных организмов типа червей, мух и улиток. Чтобы выживать в своей среде обитания, эти существа используют молекулы тех же веществ, которые используем мы для управления своей повседневной жизнью, приспосабливания к окружающей среде, испытывания всего спектра эмоций, желаний, мышления.
Кроме химических синапсов, использующих медиаторы для взаимодействия, существуют менее распространенные электрические синапсы. Их структура и функции существенно отличаются от химических и будут кратко описаны в конце главы. До начала этой части главы слово «синапс» будет употребляться в значении «химический синапс».

До начала XX века нейробиологи считали, что нервные клетки обмениваются исключительно электрическими сигналами, но даже тогда были сомнения в возможности электрического взаимодействия между нейронами из-за предположения Рамона Кахаля о существовании синаптической щели. Исследования английского ученого Джона Лэнгли (John Langley) в 1901 году подтвердили химическую связь между клетками нервных цепей. Он изучал иннервацию внутренних органов и обнаружил, что повысить артериальное давление и частоту сокращений сердца можно двумя путями:
- прямая электрическая стимуляция соответствующих нервных волокон
- инъекция экстрактов надпочечника (то есть воздействие на нервные клетки химическими веществами типа адреналина). Было доказано, что их попадание на поверхность нейронов вызывало электрические импульсы.
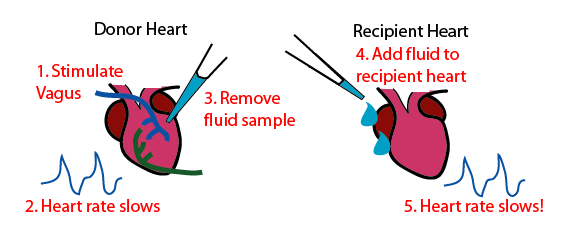
Но окончательный вердикт был вынесен в 1921 году по итогам опытов немецкого ученого Отто Лëви (Otto Loewi). Он извлек из лягушек два бьющихся сердца, одно из которых не было иннервировано, а к другому оставался присоединенным блуждающий нерв. Сердца были помещены в отдельные емкости с физиологическим раствором. Сначала с помощью электрической стимуляции блуждающего нерва Лëви заставил одно из сердец биться медленнее. После этого извлекли часть жидкости, окружающей это сердце, и добавили в емкость со вторым сердцем, которое сразу же после этого в свою очередь замедлило частоту ударов. Это стало доказательством того, что какое-то растворенное в жидкости вещество, выделенное при стимуляции блуждающим нервом, привело к замедлению сердечного ритма. А значит, сигнал, передаваемый нейроном мышце, имеет химическую природу.
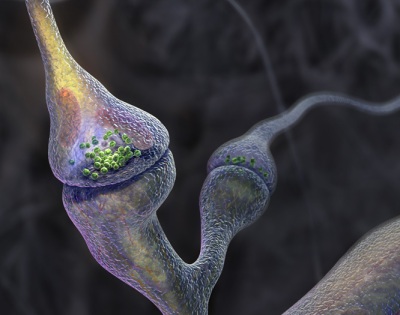
Итак, синапс — это место, где аксон одной клетки почти касается тела или дендрита другой, образуя узкую щель. Эту щель и должны преодолеть испускаемые передающим нейроном медиаторы, чтобы быть принятыми рецепторами получающей сигнал клетки.
На картинке справа изображен синапс, образованный двумя отростками разных клеток. Если рассмотреть его подробнее (на картинке внизу), то становится очевидным отличие пресинапса (части аксона, передающей сигнал) от постсинапса (части принимающей сигнал клетки): в пресинапсе есть синаптические пузырьки, или везикулы, содержащие молекулы медиатора.
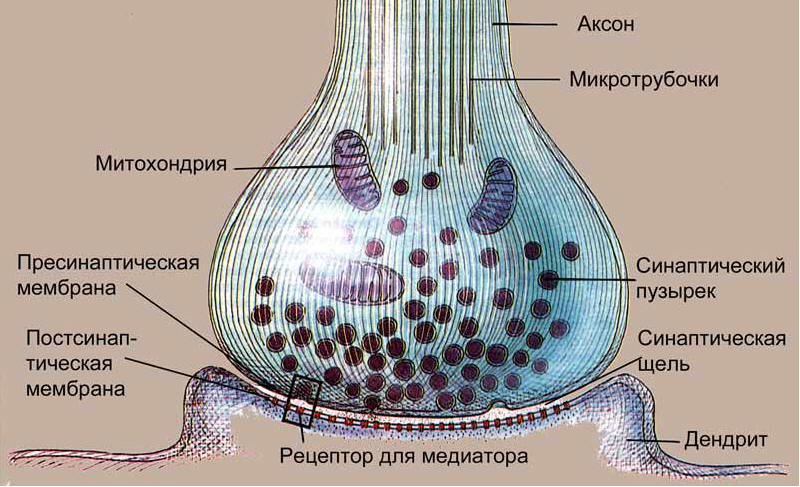
Кроме того, для обеспечения энергией процессов, происходящих в пресинаптическом окончании, в нем находится большое количество митохондрий, ведь гораздо удобнее производить энергию на месте, чем передавать откуда-то издалека. В крупных пресинапсах их может быть несколько десятков. Кроме своей основной функции, митохондрии принимают участие в синтезе и утилизации многих видов медиаторов, а также служат резервуаром для ионов кальция.
Содержимое везикул может быть разным: сейчас открыто более 100 видов медиаторов, для многих из которых все еще не установлено, какую именно роль они играют в передаче нервных импульсов. Именно медиатор определяет, какую функцию выполняет синапс, то есть какое воздействие пресинапс оказывает на постсинаптическую клетку.

Синаптические пузырьки собираются в аппарате Гольджи (смотри картинку выше), и на момент сборки в них отсутствует медиатор. Зато в их стенки сразу же встраивается фермент аденозинтрифосфатаза (АТФаза), который служит для закачки в пузырьки медиатора, а также для его выделения. Но пузырьки не отправляются в окончание аксона пустыми: в них содержатся ионы кальция, которые играют важную роль в функционировании синапса.
С помощью системы быстрого аксонного транспорта (его скорость доходит до 400 мм в сутки!) везикулы доставляются в пресинапс, где АТФаза перемещает внутрь них молекулы медиатора, находящиеся в цитоплазме пресинапса. В пузырьках медиаторы ждут своего часа: как только проходящий по аксону электрический импульс доходит до его окончаний, деполяризация мембраны этого окончания приводит к открытию потенциал-зависимых кальциевых каналов, ионы кальция входят в аксон и запускают процесс слияния везикул с клеточной мембраной пресинапса. Сливаясь, пузырьки «выворачиваются», выпуская свое содержимое в синаптическую щель. Для выброса содержимого одного пузырька требуется примерно 4 иона кальция. В течение короткого (несколько микросекунд) времени до-импульсовая концентрация ионов кальция восстанавливается, и выпуск медиатора прекращается.

Ширина синаптической щели составляет 10-50 нм, она заполнена межклеточной жидкостью. Пройдя через нее, медиатор попадает на постсинаптическое окончание, точнее — на его рецепторы. Дальше этих рецепторов медиатор не проходит, потому что его функция заключается в активации мембраны, и для этого достаточно провзаимодействовать с рецептором. На рисунке справа медиаторы изображены в виде пирамид, присоединяющихся к рецепторам постсинапса. После присоединения медиатора рецептор меняет свою конфигурацию, запуская таким образом какой-нибудь процесс в постсинаптической мембране. Например, как на схеме, расположенной ниже, открывая натрий-калиевые каналы. Их активность приводит к изменению электрического потенциала постсинапса и части окружающей его мембраны принимающей сигнал клетки.
В момент, когда клетка получит достаточное количество таких сигналов от разных постсинапсов, она перейдет порог возбуждения: будет достигнуто определенное значение мембранного потенциала, которое запускает электрический потенциал действия. И тогда по аксону принявшего сигналы нейрона пройдет электрический импульс, запустив на его концах химическую передачу этого сигнала следующим в цепи клеткам.
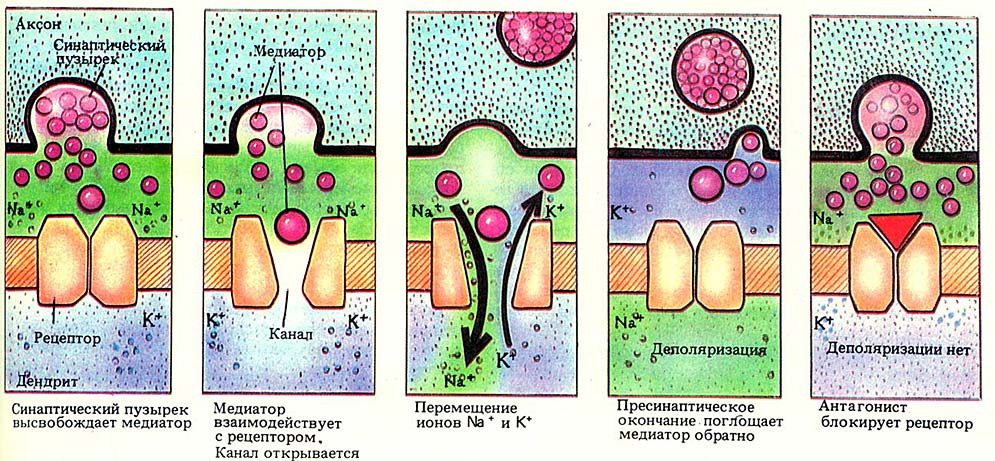
После того, как мавр медиатор сделал свое дело, он может удалиться, и часто это осуществляется наиболее экономичным путем: возвращением его в пресинаптическое окончание, где он будет помещен в везикулы и использован для передачи следующих сигналов. Процесс его возвращения будет рассмотрен ниже.

Именно на этапе попадания медиатора на рецептор процесс передачи нервного импульса по цепи может быть прерван, например присутствием антагониста (смотри пятую картинку на схеме выше), занимающего место медиатора на рецепторе, но не приводящего к нормальной реакции постсинаптической мембраны на медиатор (например, к открытию канала).
Примером такого антагониста является кофеин, который прикрепляется к рецепторам, на которые в норме присоединяется один из медиаторов (аденозин), снижающий возбуждение в нейронах головного мозга. В итоге снижающие возбуждение сигналы просто не проходят по нервным цепям и не запускают процессы, являющиеся здоровой реакцией, скажем, на усталость организма: сонливость, желание отдохнуть. При длительном воздействии кофеина организм пытается бороться с таким нарушением его сбалансированной настройки, выращивая в нейронах дополнительные рецепторы для аденозина, чтобы хотя бы часть тормозящих сигналов проходила по нервным цепям, так что постепенно кофеин перестает оказывать такое «бодрящее» действие на человека. При внезапном прекращении поступления кофеина аденозин занимает все доступные рецепторы, что может привести к усилению торможения нервной системы, проявляющемуся в утомлении, сонливости и даже депрессии.
Обычно в синапсах вырабатывается один медиатор, или один основной и один вспомогательный. Предполагается что у некоторых синапсов может выделяться и большее количество медиаторов. Общим свойством всех синапсов является то, что вырабатываемый медиатор (медиаторы) остается одним и тем же на протяжении всей жизни нейрона.
Действие разных медиаторов приводит к разным реакциям постсинаптической клетки. Можно условно разделить их на два типа: в первом случае в реакции участвует только мембрана и встроенные в нее протеины, во втором — внутриклеточные ферменты и даже ДНК.
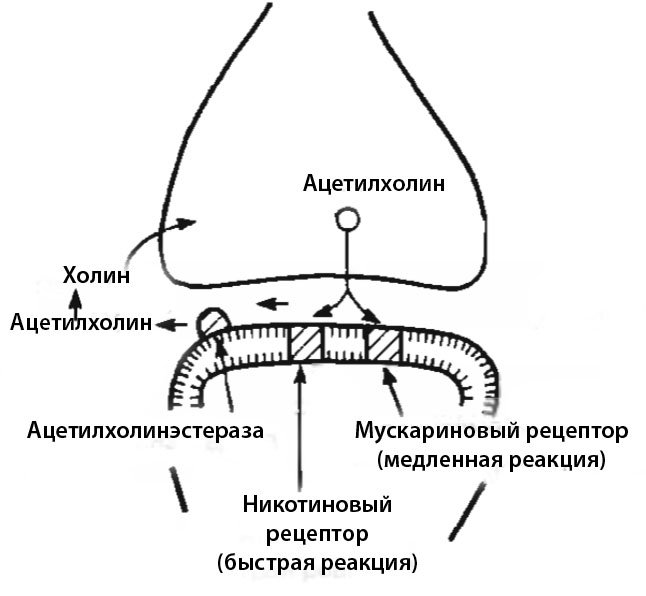
1. Примером медиаторов, передающих возбуждающий сигнал и запускающих постсинаптические реакции первого типа, является ацетилхолин. Он может связываться с двумя видами рецепторов, один из которых приводит к быстрой, но кратковременной реакции, а второй — к медленной и длительной. После связывания ацетилхолина с рецептором мембрана постсинапса резко увеличивает свою проницаемость для ионов натрия, что меняет ее электрический потенциал.
Медиатор должен быть удален сразу же после получения переданного им сигнала. За это отвечает фермент ацетилхолинэстераза, вытесняющая его с рецептора в синаптическую щель. Находящиеся в ней молекулы ацетилхолина разрушаются ферментами с образованием холина, который втягивается в пресинаптическое окончание для создания новой порции медиатора.
В тормозном синапсе первого типа молекулы медиатора, взаимодействуя с рецепторами постсинаптической мембраны, вызывают открытие каналов, позволяющих войти в клетку отрицательно заряженным ионам хлора и выйти из нее положительно заряженным ионам калия. Вход в клетку Cl– и дополнительная утечка из клетки К+ приводят к гиперполяризации постсинаптической мембраны, которую называют тормозным постсинаптическим потенциалом. Почему гиперполяризация называется тормозным потенциалом? Что и почему она тормозит? Это несложно понять: возникшая гиперполяризация, во-первых, оказывает тормозящий эффект хотя бы лишь потому, что снижает возбудимость клетки, ведь чтобы хотя бы частично гиперполяризированная клетка перешла тот порог, за которым возникнет потенциал действия, теперь необходимо сначала устранить эту гиперполяризацию, привести тем самым мембрану в состояние покоя, и уже только после этого продолжать воздействовать на нее возбуждающими импульсами. А во-вторых, тормозной постсинаптический потенциал может нейтрализовать возникший в другом месте клетки возбуждающий потенциал (подробнее про суммирование нейроном поступающих в него возбуждающих и тормозных потенциалов смотри в главе 24).

2. Медиаторы, запускающие постсинаптические реакции второго типа, также связываются с рецептором, как ацетилхолин, после чего извлекаются оттуда с возвращением в пресинапс. Но запускаемые в постсинапсе процессы существенно отличаются от тех, что возникают от действия ацетилхолина:
- рецепторный фермент активирует внутриклеточный фермент протеинкиназу
- протеинкиназа выполняет свою функцию фосфорилирования (то есть присоединения остатка фосфорной кислоты) определенных протеинов
- фосфорилированные протеины меняют проницаемость мембраны постсинапса (это совпадает с действием других медиаторов). То есть одно из отличий постсинаптической реакции второго типа заключается в присутствии внутриклеточных посредников (протеинкиназы и обработанных ей протеинов), которые запускают ответную реакцию клеточной мембраны на медиатор.
- вторым существенным отличием от реакций первого типа является то, что в реакцию второго типа может быть вовлечен и геном постсинаптической клетки, что приводит уже к долговременному изменению ее метаболизма.
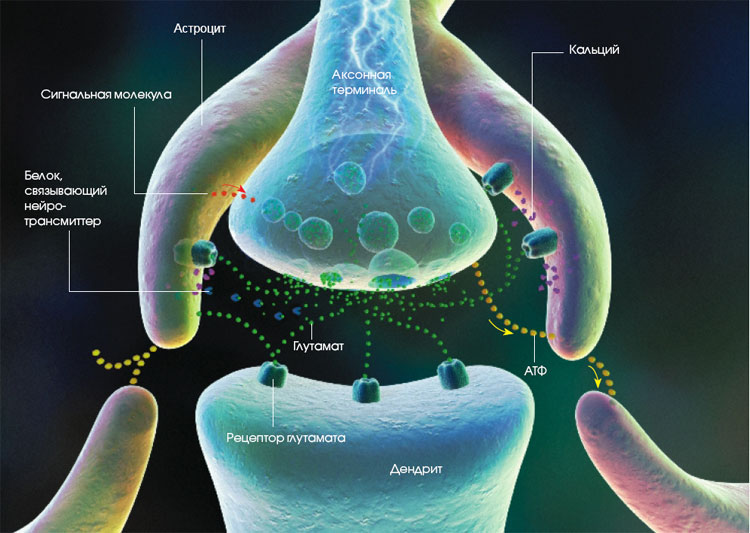
Важным участником процессов, происходящих в синапсах, являются клетки нейроглии — астроциты и олигодендроциты. Они могут активно захватывать из синаптической щели медиаторы или их составные части после того, как сигнал был передан в постсинапс, после чего передают его в пресинаптическое окончание. На картинке слева от этого абзаца — пример такой работы астроцита по возвращению глутамата в пресинапс.
Подводя итог, можно разбить процесс передачи сигнала химическим синапсом на 9 этапов:
- электрический потенциал действия доходит по аксону до пресинаптического окончания
- возникает деполяризация мембраны пресинапса, что делает ее проницаемой для ионов кальция
- ионы кальция проходят в пресинапс из межклеточного пространства, таким образом их концентрация в пресинапсе резко увеличивается
- чувствительные к кальцию протеины, встроенные в мембраны везикул, активируются высокой концентрацией ионов кальция
- эти протеины меняют свою конфигурацию, что приводит к слиянию мембран везикул с клеточной мембраной пресинапса и выбрасыванию медиатора в синаптическую щель
- медиаторы рассеиваются в пространстве щели, и часть их попадает на рецепторы постсинапса
- присоединение медиатора к рецептору приводит к изменениям в мембране постсинапса (характер изменения зависит и от вида медиатора, и от рецептора)
- изменения в мембране приводят к повышению ее проницаемости для ионов, находящихся рядом с ней, и за счет перемещения ионов с одной стороны мембраны на другую происходит изменение ее электрического потенциала. Такие изменения в электрических потенциалах всех постсинапсов принимающего сигналы нейрона суммируются им, и результатом может быть возникновение электрического импульса (в случае превалирования возбуждающих импульсов над тормозящими), или наоборот — гиперполяризация нейрона, делающая более затратным процесс его возбуждения (в случае, когда сила тормозящих сигналов превышает силу возбуждающих)
- медиатор отсоединяется от рецепторов и извлекается из синаптической щели двумя путями: 1. возвращением его в пресинапс с последующей упаковкой в везикулы и 2. расщеплением его на составные части.

В электрическом синапсе клетки контактируют гораздо плотнее: ширина синаптической щели менее 4 нанометров, и мембраны нейронов соединяются с помощью особых протеиновых образований — коннексонов. Каждый коннексон состоит из шести протеинов — коннексинов, которые образуют канал для прохождения ионов, переносящих электрический импульс.
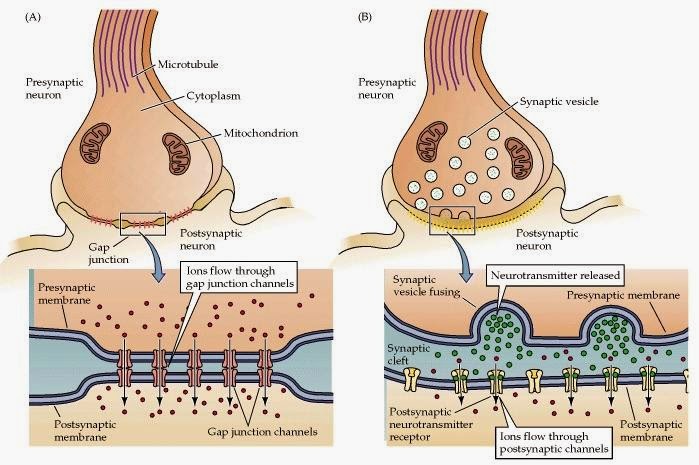
Электрический импульс проходит непосредственно от пресинаптического окончания к постсинаптическому, и отсутствие медленного химического посредника позволяет выиграть время. При передаче сигнала через химический синапс происходит задержка в 2 сотых доли секунды, а прохождение сигнала через электросинапс требует в 10 раз меньше времени. Поэтому электрические синапсы часто встречаются в тех частях нервных систем, которые отвечают за защитные рефлексы.

Та самая аплизия, чьи нейроны исследуются нейрофизиологами для поисков способов формирования памяти, используемых в том числе и мозгом человека, передает сигналы об опасности через электросинапсы.
Еще одно отличие электросинапса от химического в том, что в них возможна (и часто встречается) передача импульса в обоих направлениях.
Существенные минусы электросинапса:
- потеря силы сигнала из-за сопротивления липидосодержащих мембран обеих частей синапса
- отсутствие тех возможностей регулирования сигнала (в том числе преобразования возбуждающего сигнала в тормозящий), которые присущи химическим синапсам. Именно эти возможности позволяют создавать сложные нервные цепи, присущие высокоразвитым животным, поэтому электрические синапсы чаще встречаются у более примитивных существ.