




















Передача сигнала в нейронных цепях, о которых говорилось в предыдущей главе цитологической ветки, заключается в попеременной передаче импульса между клетками и передаче импульса внутри тела и отростков отдельно взятых нейронов. Схематично это можно представить так: сигнал возникает в рецепторах одного нейрона, проходит через него, передается другому нейрону, через тело которого в свою очередь сигнал должен пройти, чтоб быть переданным третьему, и так далее. Способы передачи сигнала внутри нейрона и между нейронами отличаются друг от друга, имеют разную природу. Эта глава целиком посвящена общему описанию передачи сигнала внутри нейрона, не затрагивая вопрос о том, как нейрон получает этот сигнал и как передает другим клеткам. Информацию о строении нейрона можно освежить, пролистав главу 20.
Чтобы провести сигнал от предшествующей клетки до последующей, нейрон генерирует электрические сигналы внутри себя. Твои движения глазами при чтении этого абзаца, ощущение мягкого кресла под попой, восприятие музыки из наушников и многое другое основаны на том, что внутри тебя проходят сотни миллиардов электрических сигналов. Такой сигнал может зародиться в спинном мозге и пройти до кончика пальца ноги по длинному аксону. Или может преодолеть ничтожно малое расстояние в глубинах мозга, ограничиваясь пределами интернейрона с короткими отростками. Любой нейрон, получивший сигнал, прогоняет его через свое тело и выросты, и этот сигнал имеет электрическую природу.
Еще в 1859 году ученые смогли измерить скорость, с которой передаются эти электрические сигналы. Оказалось, что электричество, передаваемое по живому аксону, принципиально отличается от электрического тока в металлах. По металлическому проводу электрический сигнал передается со скоростью, близкой к скорости света (300 000 километров в секунду), ведь в металле много свободных электронов. Однако, несмотря на эту скорость, сигнал ощутимо ослабевает, преодолевая большие расстояния. Если бы по аксонам сигналы передавались тем же способом, которым передаются в металлах, то нервный импульс, идущий от нервного окончания в коже большого пальца твоей ноги, полностью затухал бы, не достигая твоего мозга — электрическое сопротивление органической материи слишком велико, а сигнал слишком слаб.
Исследования показали, что электричество передается по аксонам намного медленнее, чем по проводам, и что в основе этой передачи лежит неизвестный ранее механизм, в результате которого сигнал распространяется со скоростью около 30 метров в секунду. Электрические сигналы, идущие по нервам, в отличие от сигналов, идущих по проводам, не ослабевают по ходу своего движения. Причина этого в том, что нервные окончания не пропускают через себя сигнал пассивно, просто позволяя имеющимся в них заряженным частицам передавать его друг другу. Они являются в каждой своей точке активным излучателем этого сигнала, ретранслируя его, и подробное описание этого механизма потребует отдельной главы. Таким образом, пожертвовав высокой скоростью проведения нервных импульсов, за счет активной передачи сигнала нейрон получает гарантию того, что возникший в большом пальце ноги сигнал достигнет спинного мозга, нисколько не ослабев.

Чтобы наблюдать прохождение электрической волны возбуждения, или потенциала действия ( action potential [‘ækʃən pə’tenʃəl] ), в живой клетке, достаточно простого устройства: один конец тонкой металлической проволоки помещается на наружную поверхность аксона сенсорного нейрона кожи, а другой подводится к самописцу, чертящему линию вверх при усилении сигнала, и вниз — при ослаблении. Каждое прикосновение к коже вызывает один или несколько потенциалов действия. При возникновении каждого потенциала самописец рисует узкий длинный пик.
Потенциал действия сенсорного нейрона длится всего лишь около 0,001 секунды и включает две фазы: быстрого нарастания, достигающего пика, а затем почти столь же быстрого спада возбуждения, приводящего к исходному положению. И тут самописец сообщает неожиданный факт: все потенциалы действия, возникающие в одной и той же нервной клетке, примерно одинаковы. Это можно увидеть на картинке слева: все пики, нарисованные самописцем, имеют примерно одну и ту же форму и амплитуду независимо от того, насколько сильным или продолжительным было прикосновение к коже, их вызвавшее. Слабое поглаживание или ощутимый щипок будут переданы потенциалами действия одной и той же величины. Потенциал действия представляет собой постоянный сигнал, подчиняющийся принципу «все или ничего»: после превышения раздражителем некоего порогового значения возникает всегда примерно одинаковый сигнал, не больше и не меньше обычного. А если раздражитель меньше порогового значения, то сигнал вовсе не будет передаваться: например, можно так легко коснуться кожи кончиком пера, что это прикосновение не будет чувствоваться.
Принцип «все или ничего» в возникновении потенциала действия вызывает новые вопросы. Как сенсорный нейрон сообщает о силе раздражителя — сильное или слабое давление, яркий или тусклый свет? Как он сообщает о продолжительности действия раздражителя? Наконец, как нейроны отличают один тип сенсорной информации от другого — например, как они отличают прикосновение от боли, света, запаха или звука? И как они отличают сенсорную информацию для восприятия от моторной информации для действия?
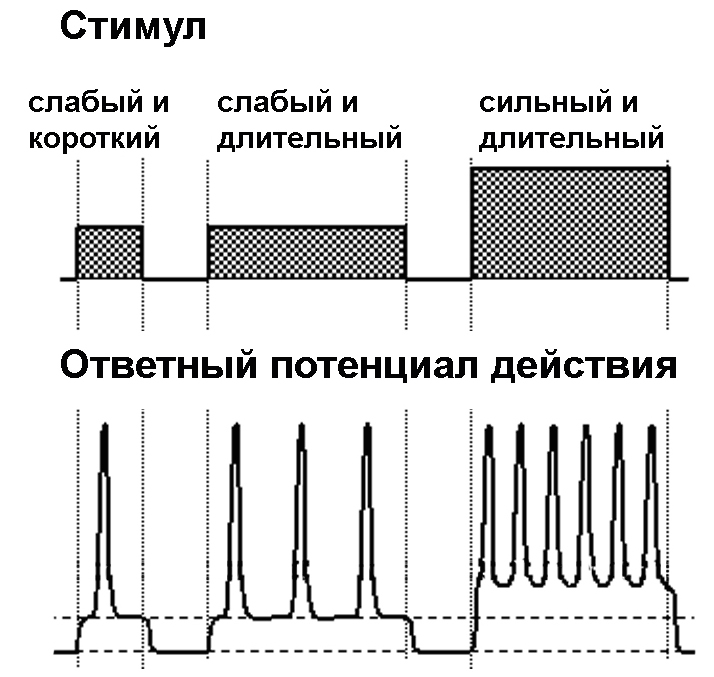
Эволюция решила вопрос о том, как сообщить о силе раздражителя, с помощью использования одного и того же вида сигналов одной и той же величины: эта сила определяется частотой ( frequency [‘friːkwənsɪ] ), с которой испускаются потенциалы действия. Слабый раздражитель, например легкое прикосновение к руке, приводит к испусканию всего двух-трех потенциалов действия в секунду, в то время как сильное давление, как при щипке или ударе по локтю, может вызвать очередь из сотни потенциалов действия в секунду. При этом продолжительность ощущения определяется продолжительностью возникновения потенциалов действия.
Используют ли нейроны разные электрические коды, сообщая мозгу, что несут информацию о разных раздражителях, таких как боль, свет или звук? Оказалось, что нет! Это удивительно, но между потенциалами действия, генерируемыми нейронами из различных сенсорных систем (например, зрительной или тактильной), разница весьма незначительна! Таким образом, характер и природа ощущения не зависят от различий в потенциалах действия (что открывает довольно захватывающую перспективу для размышлений на тему «матрицы» из одноименного фильма). Нейрон, передающий слуховую информацию, устроен точно так же, как нейрон из зрительной нервной цепи, и проводят они одни и те же потенциалы действия, одним и тем же способом. Без знания, к какой нервной цепи принадлежит конкретный нейрон, только по анализу его функционирования невозможно определить, какую информацию он несет.
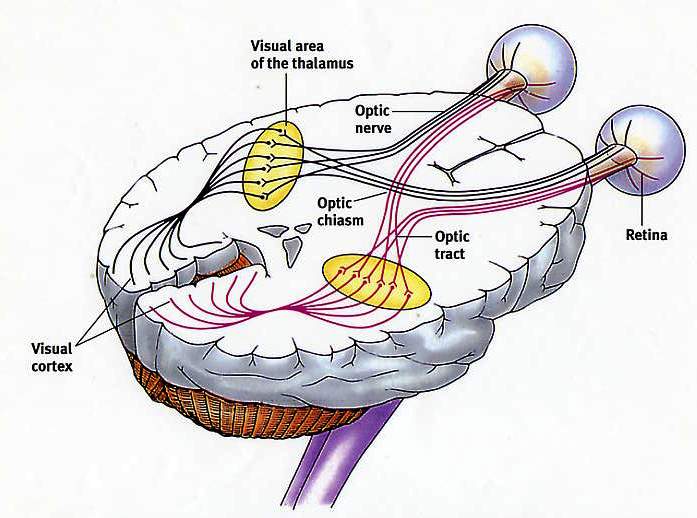
Природа передаваемой информации зависит прежде всего от типа возбуждаемых нервных волокон и специфических систем мозга, с которыми эти волокна связаны. Ощущения каждого типа передаются по своим проводящим путям, и разновидность передаваемой нейроном информации зависит именно от пути, в состав которого входит этот нейрон. В любом сенсорном проводящем пути информация передается от первого сенсорного нейрона (рецептора, реагирующего на внешний раздражитель, например прикосновение, запах или свет) к специализированным нейронам в спинном или головном мозге. Таким образом, зрительная информация отличается от слуховой лишь тем, что передается по другим проводящим путям, начинающимся в сетчатке глаза и заканчивающимся в участке мозга, который отвечает за визуальное восприятие.
Сигналы, посылаемые от моторных нейронов мозга к мышцам, также почти идентичны передаваемым по сенсорным нейронам от кожи в мозг. Они подчиняются тому же принципу «все или ничего», так же передают интенсивность сигнала с помощью частоты потенциалов действия, и так же результат сигнала зависит только от того, в какую нервную цепь включен этот нейрон. Таким образом, быстрая череда потенциалов действия, идущая по определенному проводящему пути, вызывает именно движение твоих пальцев, а не, скажем, восприятие разноцветных огней, лишь потому, что данный путь связан с мышцами рук, а не с сетчаткой глаз.

Универсальность потенциалов действия не ограничивается схожестью их проявления в разных нейронах, находящихся в пределах одного организма. Они настолько одинаковы у разных животных, что даже умудренный опытом исследователь не способен точно отличить запись потенциала действия нервного волокна кита, мыши, обезьяны или его научного руководителя. Тем не менее потенциалы действия в разных клетках не являются идентичными: небольшая разница в их амплитуде и длительности все же есть, и утверждение «все потенциалы действия одинаковы» так же неточно, как и «все бугенвиллии одинаковы».
Итак, каждый нейрон передает сигнал через свое тело и отростки одним и тем же образом. Все разнообразие информации, получаемой нами от сенсорных нейронов, все движения, которые может совершать наше тело — результат передачи единственного типа сигналов внутри нейронов. Осталась «мелочь»: понять, что же это за сигнал и как он передается.

Мы привычно отделяем все, что считаем живой природой, в том числе и себя самих, от «неживых» вещей, в том числе металлов и передающегося через них электрического тока. Тем удивительнее осознавать, что в наших телах металлы не просто присутствуют — они необходимы, без них тело не сможет существовать. Электрический ток — явление не разовое, а непрерывно возникающее в сотне миллиардов нейронов, пронизавших своими отростками все наше тело. Прямо сейчас ты можешь ощутить самые разные признаки его присутствия: то, что ты осознаешь этот текст, есть результат бесчисленных передач электрического тока. Чувство голода и удовольствие от запаха готовящейся еды, само восприятие этого запаха, прикосновение залетевшего в окно ветра к твоей коже… Перечислять можно бесконечно. И желание понять, каким же образом все это происходит, также складывается из возникающих в нейронах электрических импульсов.
Так как целью этой главы является сообщение лишь самой общей информации о прохождении нервного импульса, то здесь же необходимо рассмотреть ту среду, в которой он возникает, те условия в клетке, которые делают возможным его возникновение и передачу. Поэтому стоит начать с изучения плацдарма, на котором будут развиваться события, а именно с нейрона в состоянии покоя ( dormant state [‘dɔːmənt steɪt] ).
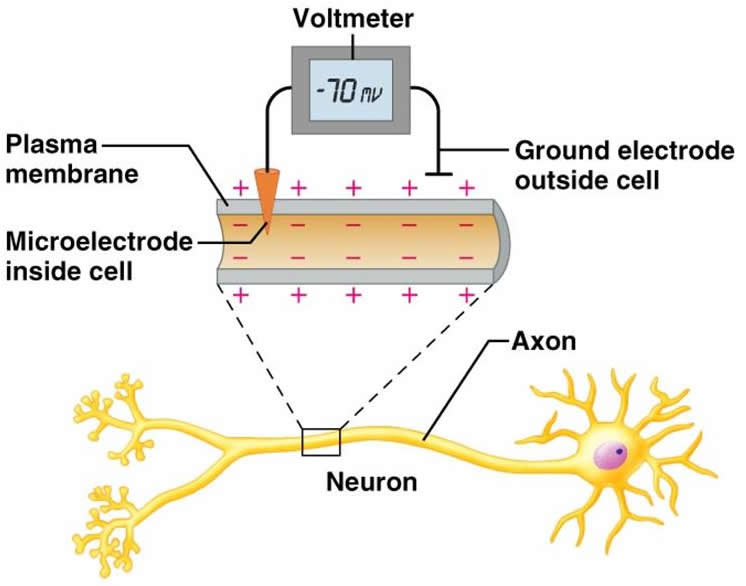
Еще в середине прошлого века ученые нашли способ установить, в какой части нейрона существует электрический заряд. Для этого используют вольтметр ( voltmeter [‘vəultˌmiːtə] ) (прибор для измерения напряжения электрического поля) с двумя электродами. Один электрод помещают внутрь нейрона, располагая его близко к клеточной мембране, а второй электрод находится в окружающей нейрон среде, с другой стороны той же мембраны. Вольтметр показывает, что с разных сторон клеточной мембраны существуют электрические заряды, отрицательный внутри клетки и положительный снаружи. Существование таких разнополюсных электрических зарядов по обе стороны мембраны создает электрическое поле, важной характеристикой которого является потенциал. Потенциал, говоря простым языком, это способность совершать работу, например работу по перетаскиванию заряженной частицы с места на место. Чем больше отрицательных зарядов накопилось по одну сторону, и чем больше положительных — по другую сторону мембраны, тем сильнее создаваемое ими электрическое поле, и тем с большей силой они способны перетаскивать туда-сюда заряженные частицы. Разницу между внешним и внутренним электрическими зарядами называют мембранным потенциалом ( membrane potential [‘membreɪn pə’tenʃəl] ) покоя. Для нейрона он равен примерно 70 мВ (милливольт), то есть 70 тысячных вольта или семь сотых вольта. Для сравнения, разность потенциалов в батарейке АА равна 1,5 вольта — в 20 раз больше. То есть мембранный потенциал покоя нейрона всего лишь в 20 раз слабее, чем между клеммами батарейки АА — довольно большой, получается. Электрический потенциал существует только на мембране, и в других своих частях нейрон электрически нейтрален.
Если написать более точно, то мембранный потенциал покоя нейрона равен -70 мВ (минус семьдесят милливольт). Знак минус означает лишь то, что отрицательный заряд находится именно внутри клетки, а не снаружи, и таким образом создаваемое электрическое поле способно перетаскивать через мембрану внутрь клетки положительно заряженные ионы.
Действующие лица в создании мембранного потенциала покоя:
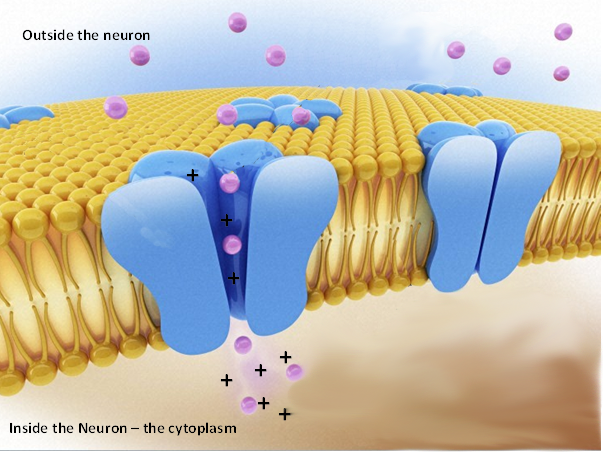
1. В клеточной мембране нейрона существуют каналы, по которым несущие электрический заряд ионы могут путешествовать сквозь нее. При этом мембрана не является всего лишь пассивной «перегородкой» между внутренней средой нейрона и окружающей его межклеточной жидкостью: специальные белки, внедренные в плоть мембраны, открывают и закрывают эти каналы, и таким образом мембрана контролирует прохождение ионов — атомов, имеющих электрический заряд. Накапливая отрицательно заряженные ионы внутри клетки, нейрон увеличивает количество отрицательных зарядов внутри, тем самым приводя к увеличению положительных зарядов снаружи, и таким образом усиливается электрический потенциал. Так как протон имеет положительный заряд, а электрон отрицательный, то при избытке протонов получается положительно заряженный ион, а при избытке электронов — отрицательно заряженный. Если хочется более подробной информации об атомах и ионах, можно вернуться в главу 4. Важно понимать, что мембранный потенциал существует именно на границе клеточной мембраны, а жидкости в целом внутри и вне нейрона остаются электрически нейтральными. Ионы, для которых мембрана проницаема, остаются вблизи нее, поскольку положительные и отрицательные заряды взаимно притягиваются друг к другу. В результате снаружи мембраны образуется слой «сидящих» на ней положительных ионов, а внутри — отрицательных. Таким образом, мембрана играет роль электрической емкости, разделяющей заряды, внутри которой есть электрическое поле. Мембрана, поэтому, является природным конденсатором.
2. отрицательно заряженные протеины, находящиеся внутри нейрона возле внутренней поверхности мембраны. Заряд протеинов всегда остается одним и тем же и является только частью общего заряда внутренней поверхности мембраны. В отличие от ионов, протеины не могут выходить из клетки и заходить в нее — для этого они слишком большие. Общий заряд меняется в зависимости от количества находящихся возле мембраны положительно заряженных ионов, концентрация которых может меняться за счет их перехода из клетки наружу, и извне вовнутрь.
3. положительно заряженные ионы калия (К+) могут свободно перемещаться между внутренней и внешней средой, когда нейрон находится в состоянии покоя. Перемещаются они через постоянно открытые проточные калиевые каналы ( flow potassium passage [fləu pə’tæsɪəm ‘pæsɪʤ] ), через которые могут пройти только ионы К+ , и ничто другое. Проточными называются каналы, не имеющие ворот, а значит открытые при любом состоянии нейрона. Внутри клетки ионов калия гораздо больше, чем снаружи. Это происходит за счет постоянной работы натрий-калиевого насоса (про него будет рассказано ниже), поэтому в состоянии покоя нейрона ионы К+ начинают перемещаться во внешнюю среду, поскольку концентрация одного и того же вещества стремится выровняться в общей системе. Если мы в бассейн с водой в одном углу выльем какое-то вещество, то его концентрация в этом углу будет очень большой, а в других частях бассейна — нулевой или очень маленькой. Однако, спустя некоторое время мы обнаружим, что концентрация этого вещества выровнялась по всему бассейну за счет броуновского движения. В этом случае говорят о «парциальном давлении» того или иного вещества, будь это жидкость или газ. Если в одном углу бассейна будет вылит спирт, то образуется большая разница в концентрации спирта между этим углом и остальным бассейном. Возникнет парциальное давление молекул спирта, и они постепенно распределятся равномерно по бассейну так, что парциальное давление исчезнет, поскольку концентрация молекул спирта везде выровняется. Таким образом, ионы К+ уносят с собой положительный заряд из нейрона, уходя наружу за счет парциального давления, которое сильнее, чем сила притяжения отрицательно заряженных протеинов, в том случае, если разница в концентрации ионов внутри и снаружи клетки достаточно велика. Так как внутри остаются отрицательно заряженные протеины, то таким образом на внутренней стороне мембраны формируется отрицательный заряд. Для ясного понимания работы клеточных механизмов важно помнить, что несмотря на постоянное вытекание ионов калия из клетки, внутри нейрона их всегда больше, чем снаружи.
4. положительно заряженные ионы натрия (Na+) находятся с внешней стороны мембраны и создают там положительный заряд. Во время фазы покоя нейрона натриевые каналы клетки закрыты, и Na+ не могут пройти внутрь, а их концентрация снаружи повышается за счет работы натрий-калиевого насоса, выводящего их из нейрона.
5. роль отрицательно заряженных ионов хлора (Cl—) и положительно заряженных ионов кальция (Ca2+) для создания мембранного потенциала невелика, поэтому их поведение пока останется за кадром.
Формирование мембранного потенциала покоя проходит в два этапа:
Этап I. создается небольшая (-10 мВ) разница потенциалов с помощью натрий-калиевого насоса.
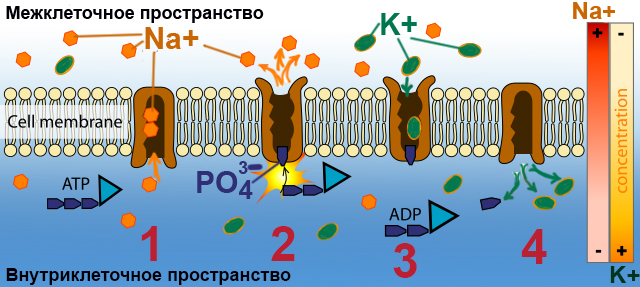
В отличие от других каналов мембраны, натрий-калиевый канал способен пропускать через себя и ионы натрия, и ионы калия. Причем Na+ может пройти сквозь него только из клетки наружу, а К+ снаружи внутрь. Один цикл работы этого канала включается в себя 4 шага:
1. «ворота» натрий-калиевого канала открыты только с внутренней стороны мембраны, и туда заходят 3 Na+
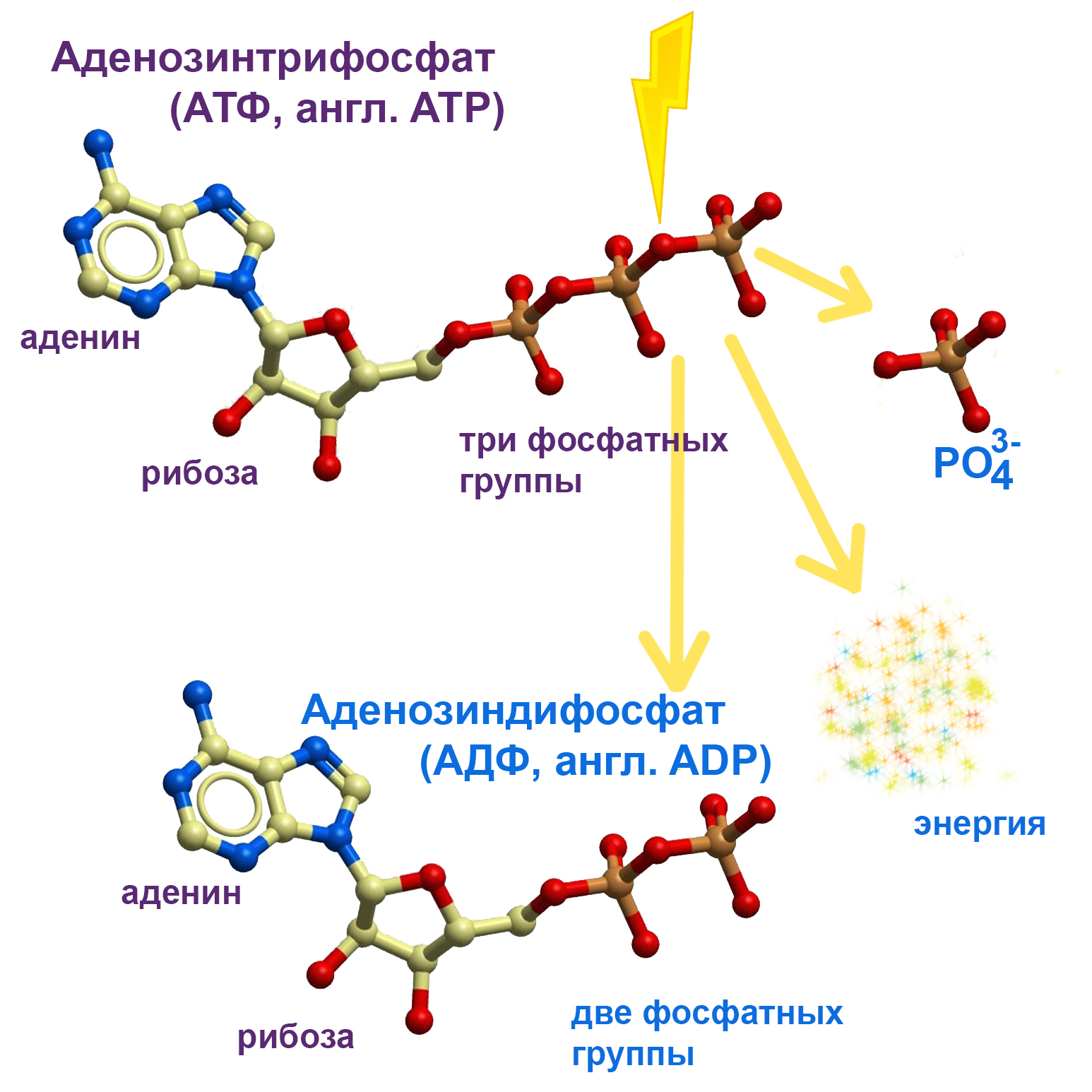
2. присутствие Na+ внутри канала воздействует на него так, что он может частично разрушить одну молекулу АТФ ( ATP ) (аденозинтрифосфата), ( adenosine triphosphate [ə’dɛnəsiːn trai’fɔsfeɪt] ) являющуюся «аккумулятором» клетки, запасающим энергию и отдающим ее при необходимости. При таком частичном разрушении, заключающемся в отщеплении от конца молекулы одной фосфатной группы PO43−, выделяется энергия, которая как раз и расходуется на перенос Na+ во внешнее пространство.
3. когда канал открывается для того, чтобы Na+ вышел наружу, он остается открытым, и в него попадают два иона К+ — их притягивают отрицательные заряды протеинов изнутри. То, что в канале, вмещающем три иона натрия, помещается всего два иона калия, вполне логично: атом калия имеет больший диаметр.
4. присутствие ионов калия теперь в свою очередь воздействует на канал так, что внешние «ворота» закрываются, а внутренние открываются, и К+ поступают во внутреннюю среду нейрона.
Таким образом работает натрий-калиевый насос, «обменивая» три иона натрия на два иона калия. Так как электрический заряд у Na+ и К+ одинаковый, получается что из клетки выводится три положительных заряда, а внутрь попадает только два. За счет этого внутренний положительный заряд клеточной мембраны снижается, а внешний — увеличивается. К тому же создается разница в концентрации Na+ и К+ по разные стороны мембраны:
=) снаружи клетки оказывается много ионов натрия, а внутри — мало. При этом натриевые каналы закрыты, и попасть назад в клетку Na+ не может, и далеко от мембраны он не уходит, так как притягивается существующим с внутренней стороны мембраны отрицательным зарядом.
=) внутри клетки много ионов калия, а вот снаружи их мало, и это приводит к вытеканию К+ из клетки через открытые во время фазы покоя нейрона калиевые каналы.
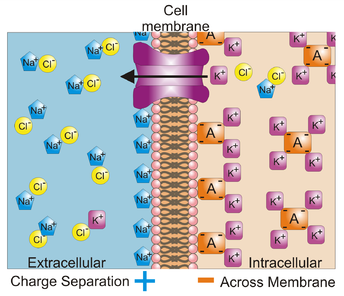
Этап II формирования мембранного потенциала покоя как раз основан на этом вытекании ионов калия из нейрона. На рисунке слева показан ионный состав мембраны в начале второго этапа формирования потенциала покоя: множество К+ и отрицательно заряженных протеинов (обозначенных А4-) внутри, и облепившие мембрану снаружи Na+. Перемещаясь во внешнюю среду, ионы калия уносят из клетки свои положительные заряды, при этом суммарный заряд внутренней мембраны снижается. Так же как положительные ионы натрия, вытекшие из клетки ионы калия остаются снаружи мембраны, притягиваемые внутренним отрицательным зарядом, и внешний положительный заряд мембраны складывается из суммы зарядов Na+ и К+. Несмотря на вытекание через проточные каналы, внутри клетки ионов калия всегда больше, чем снаружи.
Возникает вопрос: почему ионы калия не продолжают вытекать наружу до того момента, пока их количество внутри клетки и вне ее не станет одинаковым, то есть до тех пор, пока не исчезнет парциальное давление, создаваемое этими ионами? Причина этого заключается в том, что когда К+ покидают клетку, снаружи увеличивается положительный заряд, а внутри образуется избыток отрицательного заряда. Это снижает желание ионов калия выходить из клетки, ведь наружный положительный заряд их отталкивает, а внутренний отрицательный притягивает. Поэтому через какое-то время К+ перестают вытекать несмотря на то, что во внешней среде их концентрация ниже, чем во внутренней: влияние зарядов по разные стороны мембраны превышает силу парциального давления, то есть превышает стремление К+ распределиться равномерно в жидкости внутри и вне нейрона. В момент достижения этого равновесия мембранный потенциал нейрона и останавливается примерно на -70 мВ.
Как только нейроном достигнут мембранный потенциал покоя, он готов для возникновения и проведения потенциала действия, про который речь пойдет в следующей цитологической главе.
Таким образом, подытожим: неравномерность распределения ионов калия и натрия по обе стороны мембраны вызвана действием двух соперничающих сил: а) силой электрического притяжения и отталкивания, и б) силой парциального давления, возникающего при разнице в концентрациях. Работа этих двух соперничающих сил протекает в условиях существования по-разному устроенных натриевых, калиевых и натриево-калиевого каналов, которые выступают в роли регуляторов действия этих сил. Калиевый канал является проточным, то есть он всегда открыт в состоянии покоя нейрона, так что ионы К+ могут спокойно ходить туда-сюда под воздействием сил электрического отталкивания/притяжения и под воздействием силы, вызванные парциальным давлением, то есть разницей в концентрации этих ионов. Натриевый канал всегда закрыт в состоянии покоя нейрона, так что через них ионы Na+ ходить не могут. И, наконец, натриево-калиевый канал, устроенный так, что он работает как насос, который при каждом цикле выгоняет три иона натрия наружу, и загоняет два иона калия внутрь.
Вся эта конструкция и обеспечивает возникновение мембранного потенциала покоя нейрона: т.е. состояния, при котором достигается две вещи:
а) внутри есть отрицательный заряд, а снаружи — положительный.
б) внутри много ионов К+ , облепивших отрицательно заряженные части протеинов, и таким образом возникает калиевое парциальное давление — стремление ионов калия выйти наружу для выравнивания концентрации.
в) снаружи много ионов Na+, образующих отчасти пары с ионами Cl—. И таким образом возникает натриевое парциальное давление — стремление ионов натрия войти внутрь клетки для выравнивания концентрации.
В результате работы калиево-натриевого насоса мы получаем три силы, существующие на мембране: силу электрического поля и силу двух парциальных давлений. Эти силы и начинают работать, когда нейрон выходит из состояния покоя.