



















Зачем митохондриям своя ДНК? Хотя почему бы симбионтам не иметь свою ДНК в себе, производя все необходимое на месте? Зачем тогда переносить часть митохондриальной ДНК в ядро клетки, создавая необходимость транспортировки продуктов генов в митохондрии? Почему митохондрии передаются только от одного из родителей? Каким образом митохондрии, полученные от матери, уживаются с геномом клетки, составленным из ДНК матери и отца? Чем больше люди узнают о митохондриях, тем больше вопросов возникает.

Впрочем, это касается не только митохондрий: в любой области любой науки расширение сферы знаний приводит только к увеличению ее поверхности, соприкасающейся с неизвестным, вызывающим все новые вопросы, ответы на которые расширят ту самую сферу с тем же предсказуемым результатом.
Итак, ДНК современных митохондрий распределена весьма странно: небольшая часть генов содержится непосредственно в митохондриях в кольцевой хромосоме (точнее, в нескольких копиях одной и той же хромосомы в каждой митохондрии), а большая часть чертежей для производства составных частей митохондрии хранится в ядре клетки. Поэтому копирование этих генов происходит одновременно с копированием генома всего организма, а производимые по ним продукты проходят долгий путь из цитоплазмы клетки внутрь митохондрий. Тем не менее это во многом удобно: митохондрия избавлена от необходимости копировать все эти гены при размножении, считывать их и строить протеины и другие составляющие, сосредоточившись на своей главной функции по производству энергии. Зачем же тогда в митохондриях все-таки находится небольшая ДНК, для обслуживания которой требуются все эти механизмы, избавившись от которых митохондрии могли бы еще больше ресурсов бросить на основную цель их существования?
Сначала предположили, что оставшаяся в митохондриях ДНК является атавизмом, наследием поглощенной метаногеном про-митохондрии, имеющей полный бактериальный геном. В начале их симбиоза, несмотря на существование в ядре тех митохондриальных генов (м-генов), которые были необходимы для поддержания внутри метаногена комфортной для про-митохондрий среды (про это подробно написано в первой главе о митохондрии), те же самые гены хранились и в каждой из митохондрий. Про-митохондрия в начале своей жизни в качестве симбионта выглядела примерно так же, как современная бактерия на схеме слева от этого абзаца.
И очень медленно из-за невостребованности эти гены исчезали из митохондриальной хромосомы в результате самых разных мутаций. А вот клеточное ядро накапливало все больше м-генов, попадавших в цитоплазму из разрушенных симбионтов-митохондрий и встраивавшихся в геном химеры-эукариота. Как только свежевстроившийся м-ген начинал считываться, клеточные механизмы производили необходимые митохондриям продукты, освобождая симбионтов от самостоятельного их создания. А значит, митохондриальный аналог перешедшего в ядро гена больше не поддерживался в рабочем состоянии естественным отбором и стирался мутациями так же, как все предыдущие. Поэтому логично было бы предположить, что скоро и те гены, которые все еще остались в митохондриях, перейдут в ядро, что приведет к большой энергетической выгоде для эукариот: ведь из каждой митохондрии можно будет убрать громоздкие механизмы копирования, считывания и исправления ДНК, а так же все необходимое для создания протеинов.
Придя к такому выводу, ученые подсчитали, за какой срок путем естественного дрейфа из митохондрии в ядро должны были перекочевать все гены. И оказалось, что этот срок уже давно прошел. В момент появления эукариотической клетки митохондрии имели обычный бактериальный геном из нескольких тысяч генов (ученые устанавливают, каким был этот геном, изучая перенесенные в ядро м-гены у разных организмов), а сейчас митохондрии всех видов эукариот потеряли от 95 до 99,9% своих генов. Больше сотни генов в митохондриях не осталось ни у кого, но и безгеновой митохондрии тоже ни у кого не появилось. Если бы ключевую роль в этом процессе играл случай, то хотя бы несколько видов уже прошли бы путь переноса генов в ядро до конца. Но этого не произошло, и изученные на данный момент митохондрии разных видов, теряющие свои гены независимо друг от друга, сохранили один и тот же их набор, что прямо указывает на необходимость присутствия именно этих генов именно в митохондриях.
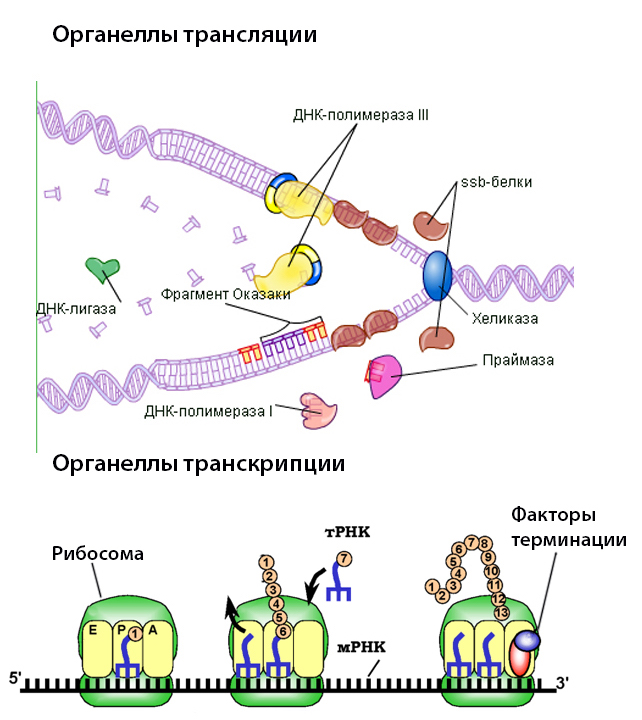
Более того, в других энергопроизводящих органеллах клеток, хлоропластах, тоже есть своя ДНК, и точно так же хлоропласты разных видов эволюционировали параллельно и независимо, оставшись каждый с одним и тем же набором генов.
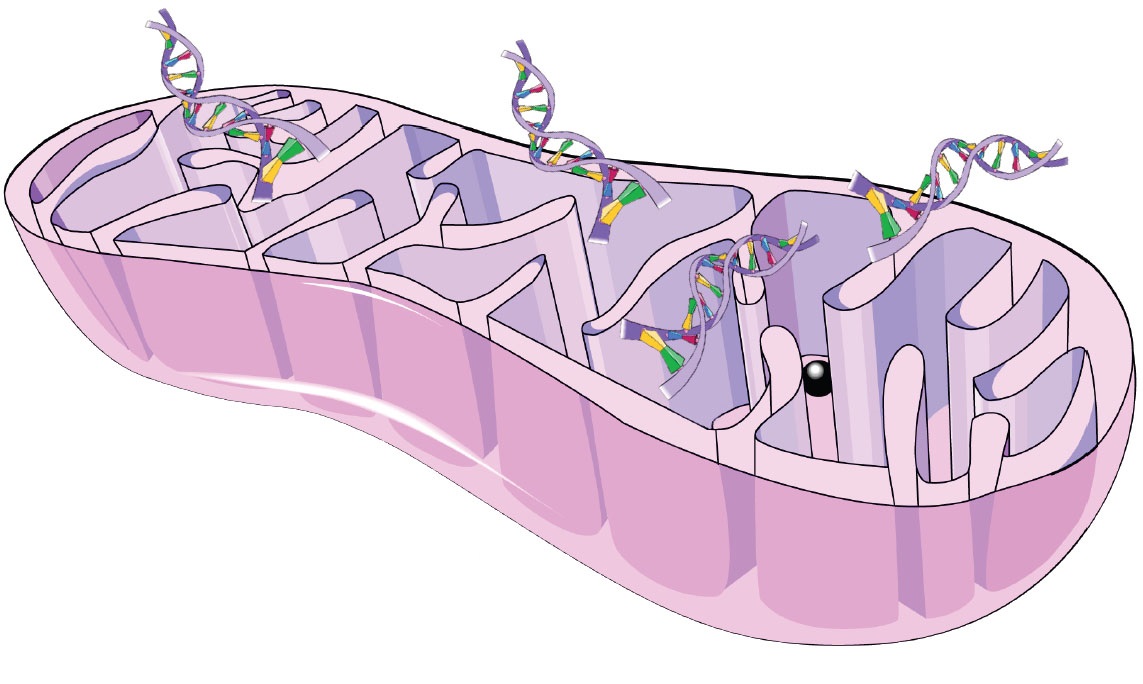
Значит, все те значительные неудобства по поддержанию собственного генома в каждой клеточной митохондрии (а в среднем в одной клетке содержится несколько сотен!) и громоздкого аппарата по его копированию-исправлению-транслированию (основные, но не все! его части ты видишь на картинке слева) чем-то перевешиваются.
И на данный момент существует непротиворечивая теория этого «чего-то»: возможность производить определенные детали митохондрии непосредственно внутри нее необходима для регулирования скорости дыхания и подстройки происходящих в митохондрии процессов под ежеминутно меняющиеся потребности всего организма.

Представь, что в одной из сотен митохондрий клетки вдруг не хватает элементов дыхательной цепи (подробно про нее смотри в предыдущей главе), или в ней недостаточно АТФ-синтаз. Она оказывается либо перегруженной пищей и кислородом и не может их достаточно быстро перерабатывать, или ее межмембранное пространство распирает от протонов, которые некуда девать — полная катастрофа в общем. Конечно же все эти отклонения от идеальной жизненной ситуации запускают множественные сигналы, направленные на выравнивание крена тонущего корабля.
Эти сигналы запускают производство именно тех деталей, которых не хватает митохондрии в данный момент, активируя считывание генов, по которым строятся протеины. Как только митохондрия будет иметь достаточно компонентов дыхательной цепи или АТФаз, «крен выровняется», сигналы о необходимости постройки новых деталей перестанут поступать, и гены опять будут выключены. Это один из удивительно элегантных в своей простоте необходимых механизмов саморегуляции клетки, малейшее его нарушение ведет к серьезной болезни или даже нежизнеспособности организма.
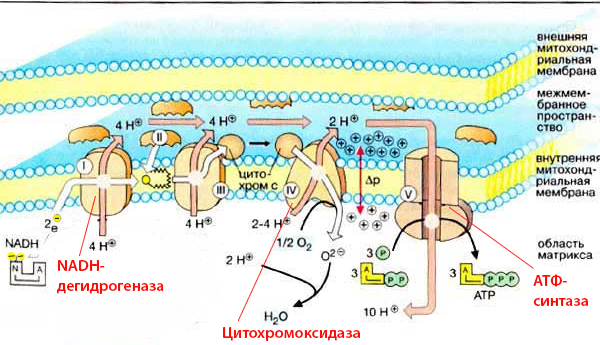
Попробуем логически определить, где должны находиться необходимые для реакции на этот сигнал бедствия гены. Представь ситуацию, что эти гены находятся в ядре клетки, содержащей пару сотен митохондрий. В одной из митохондрий возник например недостаток NADH-дегидрогеназы: первого фермента из дыхательной цепи, чья роль состоит в отрыве двух электронов от молекулы NADH, передаче их следующему ферменту и прокачке 2-4 протонов через мембрану.
На самом деле такие недостатки какого-либо фермента случаются довольно часто, ведь они периодически выходят из строя, количество потребляемой пищи постоянно меняется, потребности клетки в АТФ тоже прыгают вслед за прыжками или валяниями организма, эту клетку содержащего. Поэтому ситуация очень типичная. И вот митохондрия испускает сигнал: «нужно строить больше NADH-дегидрогеназы!», который выходит за ее пределы, проходит по цитоплазме до ядра, проникает в ядро и запускает считывание нужных генов. По клеточным меркам время прохождения этого сигнала весьма существенно, а ведь требуется еще и вытащить из ядра в цитоплазму построенную матричную РНК, создать по ней протеины, переслать их в митохондрию…
И вот тут возникает проблема гораздо более существенная, чем трата лишнего времени: при создании специализированных митохондриальных протеинов они маркируются сигналом «доставить в митохондрию», но вот в какую? Неизвестно. Поэтому в каждую из пары сотен митохондрий начинают поступать протеины, которые им не нужны. Клетка тратит ресурсы на их производство и доставку, митохондрии заполнены лишними дыхательными цепями (что приводит к неэффективности дыхательных процессов), а та единственная митохондрия, которой эти протеины нужны, не получает их в достаточном количестве, ведь ей достается в лучшем случае сотая часть произведенного. Поэтому она продолжает посылать сигналы бедствия, и хаос продолжается. Даже по этому лирико-поверхностному описанию происходящего понятно, что такая клетка нежизнеспособна. И что есть гены, которые должны считываться и транслироваться непосредственно в митохондрии, чтобы регулировать происходящие именно в ней процессы, а не полагаться на запущенный партией ядром план производства гвоздей.. то есть протеинов дыхательной цепи для всех митохондрий сразу.
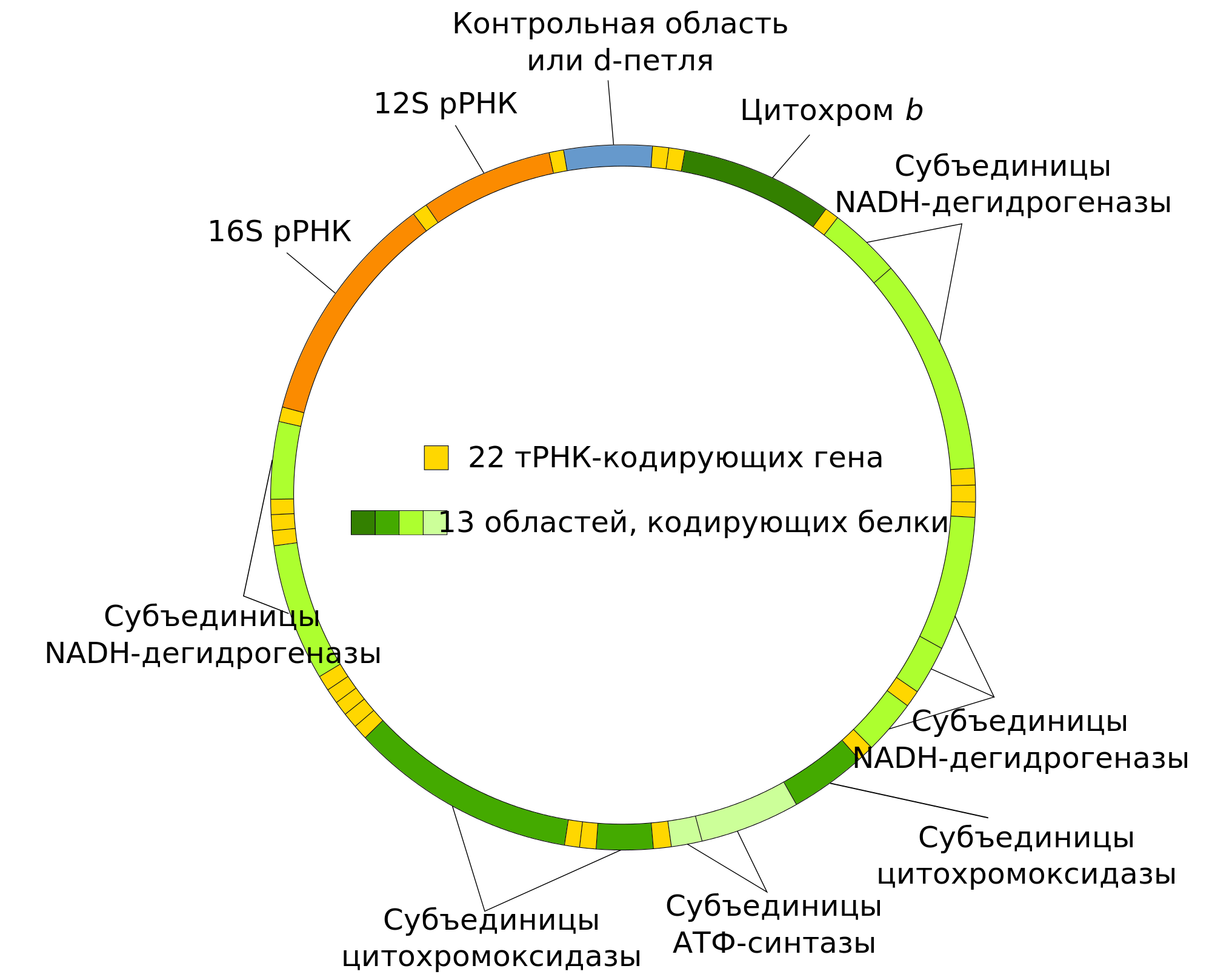
Проверив, что именно производится по оставшимся в митохондриях разных (а значит, и перемещавших м-гены в ядро независимо друг от друга) организмов, обнаружили, что это именно элементы для построения дыхательных цепей и АТФазы, а так же рибосом (то есть главной части аппарата трансляции).
Подробнее об этом (и не только) можно прочитать у Лейна в «Энергия, секс, самоубийство: митохондрия и смысл жизни». Ну и можно просто сравнить схему митохондриальной ДНК, где расшифрованы кодируемые продукты (справа от этого абзаца), со схемой дыхательной цепи (вверху), чтобы стало понятно, что именно производится в митохондрии. Конечно же, не каждый протеин, встраиваемый в эту цепь, производится на месте, часть из них строится в цитоплазме клетки. Но основные «якоря», на которые цепляются остальные детали, создаются внутри митохондрии. Что позволяет производить ровно столько ферментов, сколько нужно, и именно там, где они необходимы.
Как митохондрии связаны с сексом и как уживаются разные геномы в одной клетке, напишу в одной из следующих глав этой линии.