




















Важным свойством генетического материала является возможность его воспроизведения. Недостаточно просто иметь наследственную информацию и хранить ее: уже в момент зарождения организма, будь это деление материнской бактерии на две дочерние копии или оплодотворение яйцеклетки сперматозоидом, в результате которого вылупится мелкая черепаха, необходимо эту информацию воспроизвести. Так как хранилищем наследственной информации большинства видов живых существ является двухцепочечная молекула ДНК, эта глава посвящена именно ее копированию, или репликации ( replication [ˌreplɪ’keɪʃn] ).
Вне зависимости от размера генома клетки перед каждым ее делением этот геном должен быть полностью реплицирован, причем ровно один раз. Любое отклонение от этого правила приводит к мутации, заключающейся либо в недостатке части генетической информации в случае неполного копирования генома, либо в появлении лишних копий генов, если какая-то часть генома была скопирована дважды. Организм с недостатком генов скорее всего окажется нежизнеспособным, а вот избыток генов может быть не только лишней нагрузкой на копирующий ДНК аппарат, но и источником мутаций, как вредных, так и полезных. Например, удвоившийся ген может кодировать фермент, и клетка с двумя комплектами чертежей, по которым этот фермент производится, будет способна создавать его быстрее и в больших количествах. Для организма это может оказаться полезным, тогда его потомки с тем же набором генов будут успешнее, чем имеющие только одну копию мутировавшего гена, и в итоге полезная мутация постепенно вытеснит менее приспособленный к жизни прежний набор генов из популяции живых существ этого вида.

Возможен и другой вариант развития событий: клетка не нуждается в дополнительном выпуске фермента, который стал возможен благодаря лишней копии кодирующего его гена. Тогда использование этой копии не будет нужным для выживания организма, а значит и естественный отбор не будет поддерживать ее в первоначальном состоянии. За счет этого в ней начнут накапливаться мелкие мутации в соответствии с другими путями эволюции генов. Эти мелкие мутации будут приводить к тому, что продукт гена все сильнее будет отличаться от исходного фермента, кодируемого исходным геном. И в какой-то момент он может оказаться полезным, что повысит шансы клетки на выживание, и эта мутация закрепится таким образом в ходе естественного отбора. Такие незначительные отличия, как повышенное производство фермента или появление нового, как раз и являются основой эволюции живых существ, и могут завести очень далеко — от археоптерикса до колибри.
В случае точного, без появления такого рода мутаций копирования клетки соблюдаются два принципа:
1. Начало репликации ДНК неизбежно влечет за собой деление клетки. Клетка обычно не «передумывает» делиться и и не решает жить дальше с двумя комплектами ДНК. Так же клетка не останавливает процесс репликации до того, как весь геном будет скопирован. Раз начавшись, копирование ДНК так и идет дальше вплоть до момента получения двух идентичных ее копий.
2. Если репликация началась, то до ее завершения клетка не может разделиться. Завершение репликации ДНК является одним из сигналов, необходимых для запуска деления клетки. При делении каждая из двух дочерних клеток получает один комплект генома из двух, образовавшихся в результате копирования ДНК материнской клетки.
Процесс репликации основан на двух свойствах молекулы ДНК:
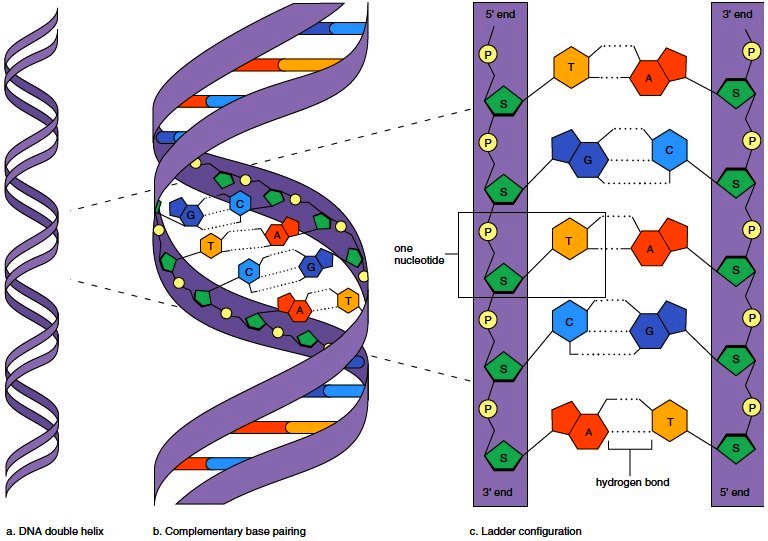
1. ДНК большинства клеток состоит из двух длинных молекул, соединенных ступеньками-нуклеотидами и закрученных в знаменитую двойную спираль. Рассмотреть ее еще раз можно в главе 5, а в 10 главе было описано то самое свойство, которое делает возможным репликацию ДНК: комплементарность.
Каждый из четырех составляющих ДНК нуклеотидов может спариться (в приличном смысле этого слова!) только с одним из этой четверки. Два спаренных нуклеотида, принадлежащие разным цепочкам, образуют одну ступеньку спирали.
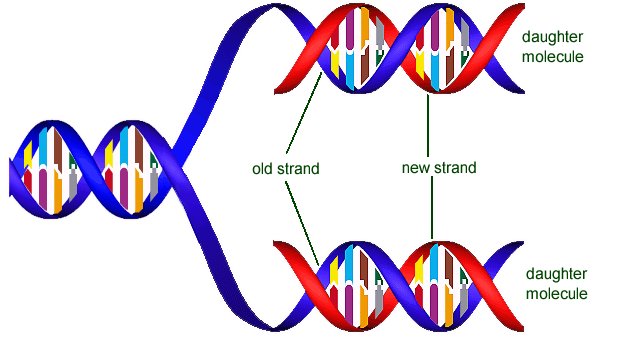
Если взять только одну нить ДНК, или матрицу, то клеточные механизмы легко достраивают к ней вторую нить, дочернюю, и делают они это согласно принципу комплементарности, т.е. сопоставляя каждому элементу матрицы комплементарный ему элемент. Аденин в материнской цепи приводит к встраиванию тимина в дочернюю цепь, а гуанин определяет встраивание туда цитозина. Так как нитей в каждой молекуле две, то не возникает проблем с поиском матриц: достаточно разделить исходную ДНК на две цепочки, и вот мы уже имеем сразу две матрицы. Теперь остается достроить к каждой из них по одной комплементарной нити, и исходная молекула ДНК превращается в две свои точные копии. Репликацию ДНК называют полуконсервативной, подчеркивая этим то, что одна из нитей этой копии принадлежала исходной ДНК.
2. Разделение молекулы ДНК на две матричных цепи основано на особенностях связей между нуклеотидами, соединяющих эти цепи — они, с одной стороны, сравнительно слабые и все же, с другой стороны, достаточно прочные. Если бы связь между нуклеотидами двух цепей была настолько же прочной, как связь между составляющими каждый нуклеотид атомами, то разделение было бы гораздо более энергозатратным и длительным процессом. Если же эта связь была бы слабее, чем она есть, то цепи ДНК слишком легко отделялись бы друг от друга, что приводило бы к повреждению наследственной информации.
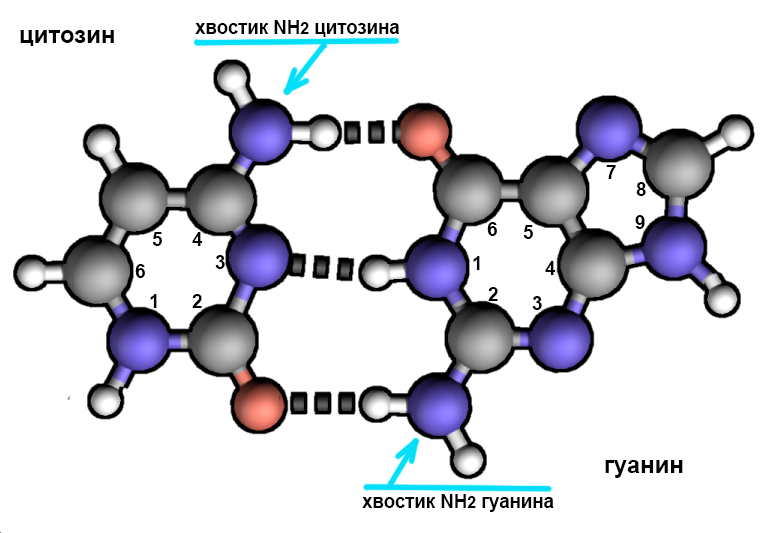
Таким образом, само существование нашей жизни зависит от использования в спирали ДНК связи оптимальной степени прочности: ее называют водородной связью, подробности о ее использовании в ДНК описаны в главе 10, а глава 15 касается (слегка поглаживая) механизма ее возникновения. На схеме справа более прочные связи между атомами нуклеотидов изображены сплошными линиями, а водородные связи между нуклеотидами пунктирные.
Для начала стоит рассмотреть процесс копирования ДНК очень поверхностно:
1. две комплементарные нити ДНК разъединяются на небольшом участке, называемом репликативной вилкой ( replicative fork [‘rɛplɪkətɪv fɔːk] )
2. из плавающих вокруг свободных нуклеотидов к каждой из расплетенных матричных цепей достраивается дочерняя цепь
3. по мере достраивания обеих матричных цепей до двойной молекулы репликативная вилка сдвигается, проходя по всей длине исходной молекулы ДНК. В результате появляются две идентичных двойных спирали. Одна из цепей каждой спирали принадлежала материнской ДНК, а вторая была собрана в процессе репликации.
Вот собственно и вся репликация: после этих трех шагов клетка готова к делению.
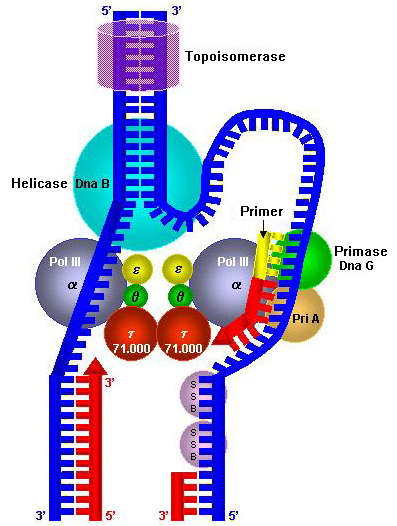
Так же как процесс создания протеина имеет свою основную рабочую единицу, рибосому, так и процесс воспроизведения ДНК выполняется реплисомой ( replisome ). Но в отличие от состоящей из двух субъединиц рибосомы реплисома представляет собой белковый комплекс, состоящий из гораздо большего числа компонентов, которые не связываются в одну органеллу, в отличие от двух рибосомальных субъединиц. Как мы помним, рибосома объединяется в одну органеллу только в момент синтеза протеина, существуя в виде отдельных субъединиц все остальное время. Так же ведет себя и реплисома. Она не существует в клетке вне процесса репликации. Этот белковый комплекс собирается непосредственно на ДНК, которую необходимо удваивать в этот момент, и разъединяется сразу же после завершения репликации. На рисунке справа изображена реплисома кишечной палочки, копирующая ее ДНК: все, что не является ДНК на этой картинке, представляет собой разные протеины реплисомы.
Если рибосомы прокариот, эукариот и органелл отличаются друг от друга незначительно, то между их реплисомами отличия уже более существенные. Один из признаков того, что реплисомы различных живых существ отличаются больше, чем их рибосомы, демонстрируют бактериофаги своими предпочтениями при размножении. Заразивший бактерию фаг пользуется ее рибосомами для построения необходимых ему белков по чертежам со своей ДНК. Но вот реплисома бактерии ему уже не подходит, и для воспроизведения своей наследственной информации он собирает свой собственный копировальный аппарат, похожий на бактериальный, но имеющий существенные отличия.
В этой главе при упоминании части реплисомы не будет конкретизации того, какой именно протеин играет эту роль у эукариот, прокариот или бактериофагов. Будет рассматриваться общая для различных организмов функция этих основных частей, а их строение и особенности функционирования останутся за кадром.
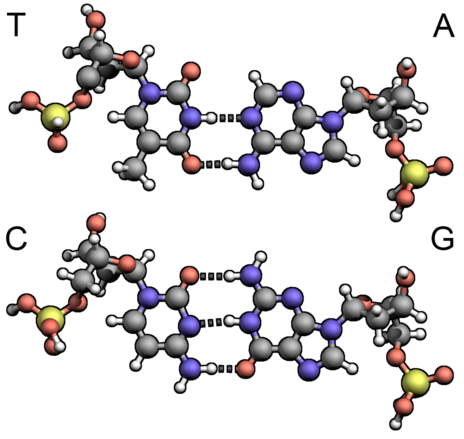
Для начала, или инициации, воспроизведения ДНК две ее цепи должны разделиться, то есть водородные связи между нуклеотидами этих цепей должны быть разрушены. Это разделение начинается в особом месте ДНК, так называемой точке начала репликации ( origin of replication [‘ɔrɪʤɪn ɔf replɪ’keɪʃn] ). Во многих русскоязычных источниках встречается заменяющая этот длинный термин калька с английского: ориджин. Структура этого фрагмента у разных организмов отличается, но общим для всех является большое количество нуклеотидных пар аденин-тимин на этом участке. Объясняется это просто: между этой парой нуклеотидов, как легко видеть, есть лишь две водородные связи, на разрывание которых требуется меньше энергии, чем на разрушение трех связей между другой парой нуклеотидов: цитозином и гуанином.
В одном ориджине могут образоваться две репликативные вилки. В этом случае они идут в противоположных направлениях, и на каждой из них собирается собственная реплисома. Независимо от того, идет репликация в обоих направлениях или только в одном, расплетенные нити матричной ДНК и строящиеся по ним дочерние образуют так называемый репликационный глазок.

Организмы с коротким геномом (например, одноклеточные) скромно обходятся минимальным количеством точек начала репликации: от одной до четырех. Геномы эукариот намного больше и требовательнее: если количество пар нуклеотидов бактерии исчисляется миллионами, то в геномах млекопитающих счет идет на миллиарды. Если бы репликация генома-миллиардера начиналась всего в четырех местах, она длилась бы слишком долго.
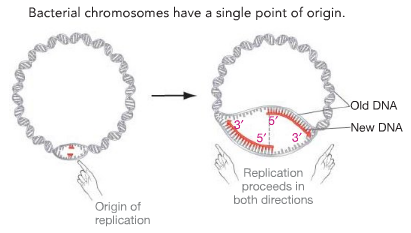
Например, геном человека содержит до ста тысяч ориджинов, поэтому в нем находится около ста тысяч репликонов ( replicon [‘rɛplɪkɒn] ) — единиц ДНК, являющихся объектом одного акта репликации. А геном бактерии, содержащий всего одну хромосому, представляет собой один-единственный репликон.
Теперь вернемся к реплисоме. Органеллы, входящие в её состав, должны выполнять множество функций: разъединять цепи ДНК, поддерживать их в таком состоянии, достраивать комплементарную цепь и т.д. Целью этой главы является получение лишь самого общего представления об этих процессах, поэтому будет достаточно рассмотреть только основных их участников.
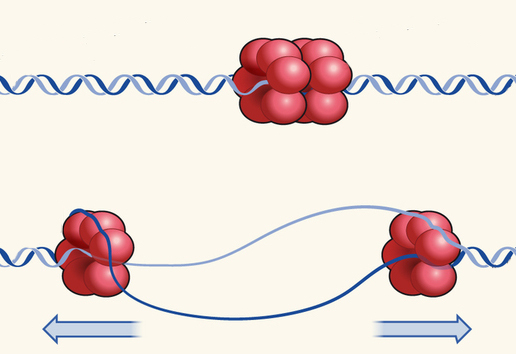
Фермент хеликаза ( helicase ), разъединяющий цепи ДНК, представляет собой кольцо из нескольких повторяющихся фрагментов. Она скользит по одной из цепей ДНК, непрерывно получая энергию от разрушения молекул АТФ и используя ее для разрыва водородных связей между цепями.

Разъединенные цепи ДНК имеют слишком много выступов, притягательных для положительно и отрицательно заряженных частиц. Среди них есть те, к которым с помощью водородных связей должны присоединиться нуклеотиды дочерних цепей, а также те, которые необходимы для сворачивания ДНК в компактную форму хромосомы. До момента, когда их участие будет необходимо, их совершенно необходимо каким-то образом закрыть, оградив от несанкционированного прилипания посторонних молекул или соседних участков самой расплетенной цепи. Этим занимаются SSB-протеины, ( SSB protein ), ( single-strand binding protein ), которые уступят свое место только ферментам, осуществляющим синтез нуклеотидной цепи. На схемах слева и справа показано, как работают хеликазы и SSB-протеины.
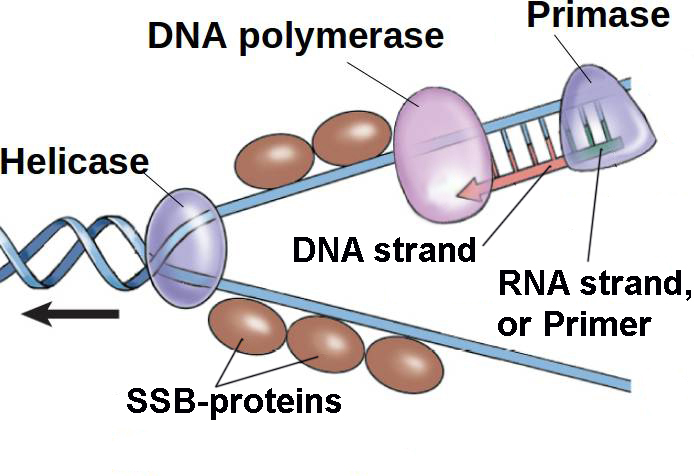
Когда хеликаза рассоединила цепи ДНК, и когда SSB-протеины заняли свое место, пора вступать в работу ферменту, который создает дочерние цепи путем присоединения комплементарных нуклеотидов. Этим ферментом является ДНК-полимераза ( DNA-polymerase ).
В процессе копирования ДНК происходит множество проверок правильности присоединения нуклеотида, которые предохраняют от присоединения неправильного нуклеотида. Это неудивительно, учитывая важность передачи наследственной информации в точности. Одним из таких предохранителей является неспособность ДНК-полимеразы начать присоединять нуклеотиды в любом месте, где ей вздумается. Она не может начать строить по матричной цепи дочернюю цепь с нуля и в произвольном месте: ей необходимо существование хотя бы небольшого отрезка дочерней цепи, к которой она будет присоединять нуклеотиды. Это кажется тупиком: если ДНК-полимераза может только достраивать уже начатую цепь, то откуда это начало будет браться?
Именно для создания такого начала цепи существует отдельный фермент: ДНК-праймаза ( DNA-primase ). Она присоединяет к матричной цепи всего-лишь 11-12 нуклеотидов, которые называют затравкой или праймером ( primer [‘praɪmə] ), а ДНК-полимераза, обнаруживая недостроенную цепь, садится на нее и достраивает, сбрасывая по ходу движения SSB-протеины со своего пути — см. картинку выше справа.
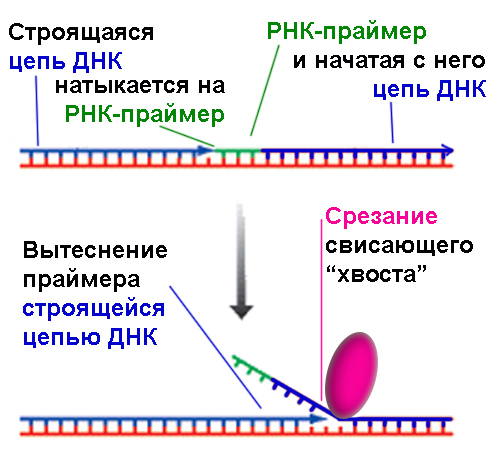
Есть еще один тонкий момент в этом процессе, который сейчас можно рассмотреть и понять. Еще одним предохранителем является создание затравки не из дезоксирибонуклеотидов, строительных единиц ДНК, а из рибонуклеотидов, являющихся «кирпичиками» РНК. То есть затравка представляет собой участок РНК, вклинившийся в строящуюся цепь ДНК. Как уже упоминалось в главе 16, рибонуклеотиды могут спариваться с нуклеотидами цепи ДНК так же успешно, как и дезоксирибонуклеотиды друг с другом. В чем может быть эволюционная выгодность такого механизма? Дело в том, что при присоединении самых первых нуклеотидов вероятность ошибки больше, и поэтому затравка создается из отличающихся «кирпичиков», чтобы потом распознать их и вырезать. Процесс этого вырезания требует участия нескольких ферментов, и в этой главе детально рассмотрен не будет. Посмотрим на схему слева. В верхней части рисунка мы видим — что происходит на стыке двух соседних репликонов. ДНК-полимераза делает свое дело, и вдруг натыкается на РНК-праймер, с которого началось построение соседнего участка ДНК. Дальше идем в нижнюю часть рисунка: ДНК-полимераза вытесняет праймер, достраивая цепь ДНК, а специальный фермент, изображенный красным овалом, срезает РНК-праймер, захватывая для надежности еще несколько достроенных за ним нуклеотидов. После этого два репликона сшиваются, и мы получаем единую цепь надстроенной нити ДНК.
Есть еще одна теория, в рамках которой можно объяснить существование РНК-затравки. Существует обоснованное предположение, что не все четыре вида молекул, образующих современные организмы, появились на Земле одновременно, начав сотрудничество, результатом которого является все разнообразие жизни. Учитывая огромное количество функций, выполняемых в клетках молекулами РНК, и их способность выполнять функции других молекул (хранить наследственную информацию, как молекула ДНК, а также играть некоторые роли, которые свойственны протеиновым ферментам), ученые выдвинули теорию «мира РНК». Предполагают, что от существ того мира, построенных из молекул РНК, современные клетки унаследовали РНК-содержащие органеллы, участвующие в частности в транскрипции и трансляции протеинов. Научившись создавать протеины, существа из «мира РНК» далее прошли сквозь естественный отбор, в результате которого оказалось, что многие клеточные функции протеиновые ферменты выполняют гораздо успешнее, чем их рибонуклеиновые аналоги. Поэтому выжили те организмы, в которых легко приспосабливаемые для выполнения различных функций протеины во многом заменили органеллы, построенные из РНК, а частично усовершенствовали их (примером такого сотрудничества протеинов и РНК является рибосома, о которой говорилось в главе 23).

В «мире РНК», как и в современном мире, существовали вирусы: существа, не имеющие своего копировального аппарата для воспроизведения наследственной информации, и стремящиеся воспользоваться теми, кто этот аппарат имеет. Древние вирусы встраивались в примитивные РНК-существ и заставляли их воспроизводить свои копии точно так же, как современные вирусы инфицируют клетки бактерий и эукариот, размножаясь с помощью их органелл. Успешнее выживали те РНК-существа, которые оказались способными бороться с вирусами, то есть те, в которых в результате мутаций возникали механизмы борьбы с вирусами. К таким механизмам относятся ферменты, которые разрезают молекулы, несущие вирусную наследственную информацию. И в какой-то момент появилась мутация вирусов, наследственная информация которых была записана не с помощью РНК, а с помощью ДНК. ДНК отлично подходили для хранения наследственной информации, и к тому же приспособленные для разрушения молекул РНК ферменты потенциальных хозяев не могли причинить такой же вред молекуле ДНК. Так появились первые вирусы, чей геном был закодирован в ДНК, а заражаемые ими существа продолжали пользоваться для хранения генома молекулами РНК. На этом этапе вирусы победили. Но естественный отбор идет непрерывно и везде в живой природе, и в каждой победе скрывается залог будущего поражения, в котором скрывается залог будущей победы и т.д.
Молекула ДНК в целом более стабильна и менее подвержена мутациям, чем молекула РНК. Поэтому неудивительно, что ДНК-вирусы процветали, и появлялись разные их виды. Один из этих вирусов сделал обычную в мире вирусов вещь, которая часто случается и сейчас: вместо того, чтоб превратить зараженного хозяина в машину по производству вирусов, он встроил свой геном в геном хозяина. Отличие от всех предыдущих и современных нам подобных встраиваний было в том, что в тот раз геном вируса был представлен молекулой ДНК, а геном хозяина — молекулой РНК. Получилось существо, геном которого состоял из смеси рибонуклеотидов и дезоксирибонуклеотидов. Бесчисленные поколения его потомков эволюционировали, находя применение для закодированных в ДНК генов, изменяя их, дублируя, опять изменяя… И постепенно вся их наследственная информация стала храниться в более стабильных молекулах ДНК. Таким образом вирусы постепенно и неизбежно передали свое открытие.
Вот так выглядит этот переход от существ, хранящих свой геном в РНК, к ДНК-геномным существам, с точки зрения гипотезы Патрика Фортера:
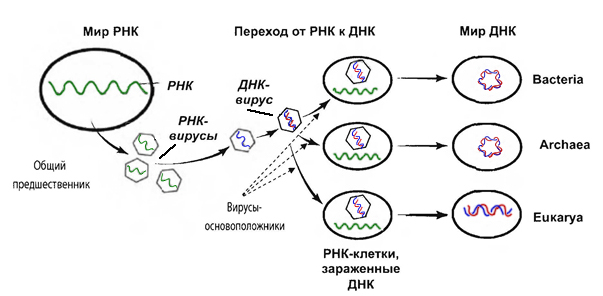 Аппарат воспроизведения генома получившихся существ был создан для копирования молекул РНК, а не ДНК. Во время и после перехода на новый носитель генома аппарат его воспроизведения менялся, и могло случиться так, что кое-что осталось прежним: при создании дочерней цепи нуклеотидов по матрице родителя сначала создается РНК-затравка. И только после того, как будут присоединены несколько рибонуклеотидов, ДНК-полимераза может начать работу по созданию цепи ДНК. Возможно, что создание коротких участков РНК на бесчисленных репликонах во время копирования твоей ДНК является отголоском того времени, когда наши предки обходились одинарной цепочкой рибонуклеиновой кислоты для хранения своего генома.
Аппарат воспроизведения генома получившихся существ был создан для копирования молекул РНК, а не ДНК. Во время и после перехода на новый носитель генома аппарат его воспроизведения менялся, и могло случиться так, что кое-что осталось прежним: при создании дочерней цепи нуклеотидов по матрице родителя сначала создается РНК-затравка. И только после того, как будут присоединены несколько рибонуклеотидов, ДНК-полимераза может начать работу по созданию цепи ДНК. Возможно, что создание коротких участков РНК на бесчисленных репликонах во время копирования твоей ДНК является отголоском того времени, когда наши предки обходились одинарной цепочкой рибонуклеиновой кислоты для хранения своего генома.
После погружения в теории о мире РНК пора вернуться в мир ДНК, к являющемуся предметом этой главы процессу репликации. Среди всех этих сложностей с расплетанием нитей ДНК в ориджине, удерживании их разъединенными, присоединением и срезанием затравок, работа ДНК-полимеразы, выполняющей основную работу по копированию ДНК, кажется очень простой: взять плавающий в цитоплазме нуклеотид, встроить его, проверить правильность встраивания и двигаться дальше. Именно этим она и занимается, проходя по всей длине репликона, пока не наткнется или на движущуюся навстречу репликативную вилку, или на участок терминации ( termination [tɜːmɪ’neɪʃn] ): особую последовательность нуклеотидов, являющуюся сигналом остановить репликацию. Стоит отметить, что прохождение ДНК-полимеразы по обеим цепям молекулы ДНК совсем не простой процесс. Настолько непростой, что для его описания потребуется целая глава, которая будет следующей в генетической ветке учебника.
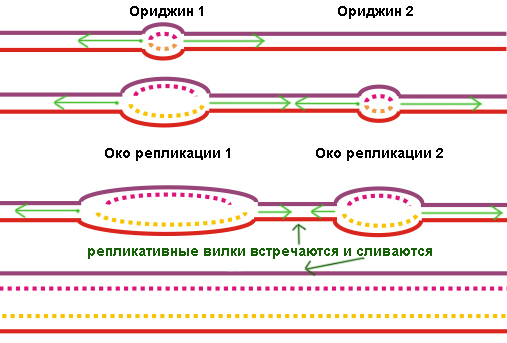
Хромосомы эукариот состоят из огромного количества репликонов, поэтому их репликативные вилки очень часто сталкиваются, двигаясь навстречу друг другу. Каждая из них достраивает свой участок ДНК, пока не натыкается на встречный конец дочерней цепи. Так как свободных нуклеотидов матричной цепи больше нет и достраивать больше нечего, реплисома покидает ДНК, оставляя позади двойную спираль, одна из цепей которой только что была синтезирована.
Итак, процесс воспроизводства ДНК происходит следующим образом:
=) в точке инициации на каждом репликоне генома цепи ДНК разъединяются
=) на разъединенных цепях собираются одна или две реплисомы, задача которых — скопировать участок ДНК и включить его в общую цепь, собранную другими реплисомами. Реплисома состоит из большого количества протеинов, выполняющих разные функции. Наиболее значимыми являются:
* расплетающая нити ДНК хеликаза
* удерживающие эти нити расплетенными SSB-протеины
* создающая РНК-затравку праймаза
* строящая дочернюю цепь ДНК-полимераза
=) завершение репликации происходит либо при слиянии репликационных вилок, расположенных на одной хромосоме, либо при попадании реплисомы на участок терминации, дающий сигнал о завершении синтеза.
Процесс репликации сейчас является объектом активного изучения, и многие его механизмы остаются не до конца понятыми или не обнаруженными. Например, до сих пор неизвестно, каким образом миллионы реплисом, копирующие огромный геном эукариот, координируют свои действия, какие клеточные механизмы дают каждой из них сигнал «пора начинать репликацию». Неизвестно, каким образом все они передают обратный сигнал о том, что репликация закончена. А этот сигнал необходим клетке для запуска деления, ведь только после завершения копирования ДНК на каждом ее участке клетка может разделиться с образованием двух идентичных копий.