



















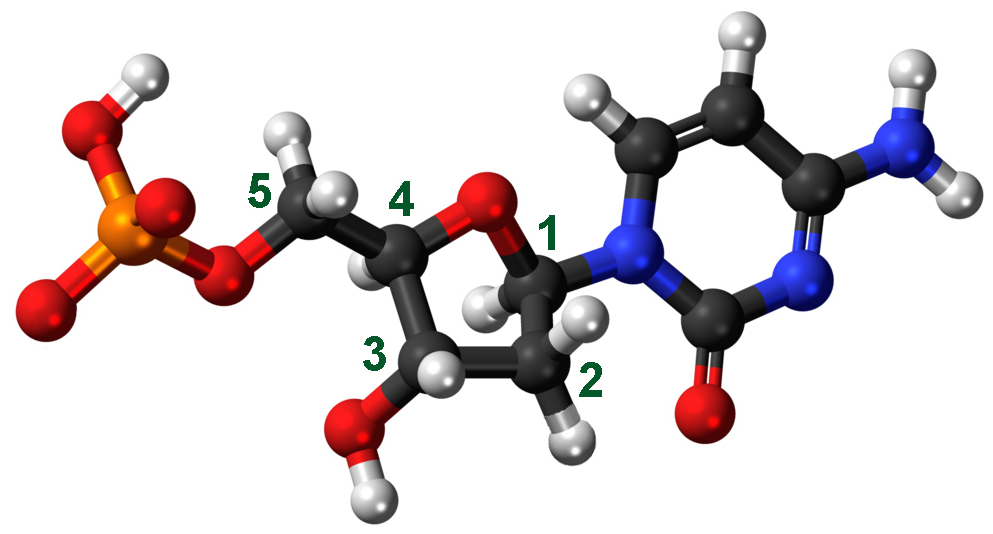
Перед подробным изучением процесса сборки полинуклеотидной (т.е. состоящей из множества нуклеотидов) цепи ДНК стоит вспомнить, что представляет собой отдельный нуклеотид. На схеме справа изображен цитозин, состоящий из:
— азотистого основания (в правой части схемы, атомы азота обозначены синим цветом), являющегося кодирующей частью нуклеотида. Именно оно отличает один нуклеотид от другого.
— сахара дезоксирибозы. Атомы углерода, образующие дезоксирибозу, пронумерованы.
— фосфатной группы (оранжево-красной слева).
Сахар и фосфат являются некодирующими частями нуклеотида, их функция заключается в создании каркаса ДНК. Именно с ними работают ДНК-полимеразы, создавая связи между нуклеотидами.
В любой клетке существует несколько видов ДНК-полимераз, различающихся по функциям на две группы:
1. способные осуществлять синтез большого фрагмента ДНК полимеразы, участвующие в репликации ДНК. Эти же ферменты используются и в процессе репарации, или исправления длинного поврежденного участка ДНК, так как они способны заново создать часть восстанавливаемой цепи. Иногда эту группу ДНК-полимераз называют ДНК-репликазами. В дальнейшем ДНК-полимеразами без уточнений будут называться исплючительно ферменты этой первой группы.
2. вспомогательные ДНК-полимеразы участвуют в процессах репликации и репарации «на вторых ролях». Некоторые из них способны восстанавливать повреждения ДНК размером в несколько нуклеотидов, другие необходимы для устранения возникающих при репликации проблем. Например, наткнувшись на повреждения матричной ДНК, репликативная вилка останавливается, и возобновляет ее движение одна из вспомогательных ДНК-полимераз. А вспомогательная ДНК-полимераза α у эукариот содержит в себе праймазу, поэтому ее задачей является создание затравки для дальнейшего синтеза цепи ДНК-полимеразой первой группы.
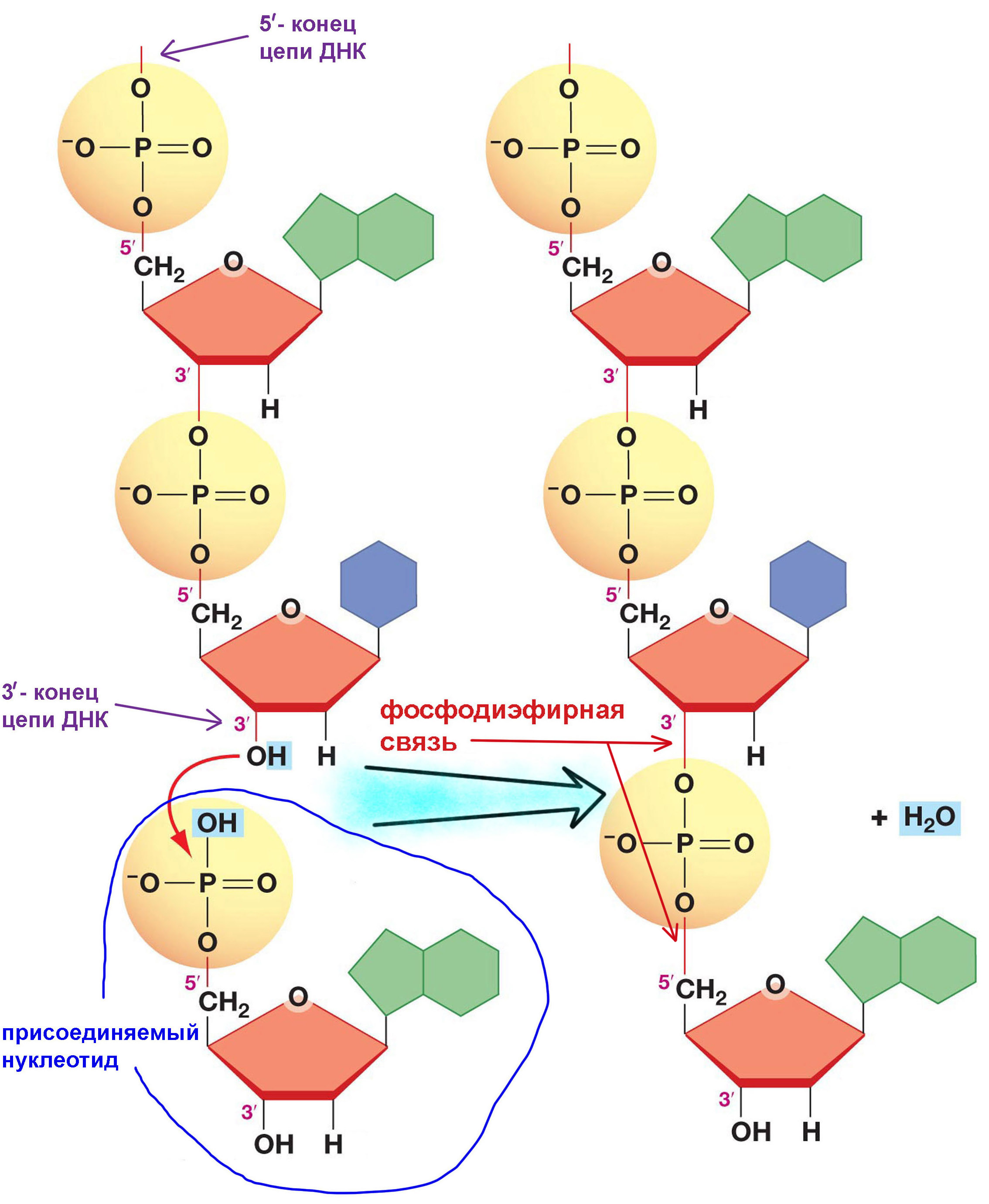
На схеме слева изображено присоединение нуклеотида, находящегося внизу картинки, к концу уже существующей цепи. При формировании полинуклеотидной цепи каждый нуклеотид прикреплен к предыдущему своей фосфатной группой, подставляя свою дезоксирибозу для присоединения следующего. Поэтому с одной стороны такая цепь заканчивается торчащей дезоксирибозой, точнее даже подвешенной к ее третьему по счету атому углерода гидроксильной группой (-ОН). Именно здесь будет присоединяться следующий нуклеотид, поэтому такой заканчивающийся свободной дезоксирибозой конец называют 3′-концом цепи ДНК. Другой конец молекулы заканчивается фосфатом последнего нуклеотида, прикрепленным к пятому атому углерода дезоксирибозы. Его обозначают 5′-конец.
Все без исключения ДНК-полимеразы не могут работать с 5′-концом цепи ДНК. Они способны присоединить новый нуклеотид только к 3′-концу. Поэтому дочерняя цепь всегда строится в направлении от её 5′-конца к 3′-концу. ДНК-полимераза соединяет атом фосфора из фосфатной группы нового «кирпичика» с атомом кислорода из дезоксирибозы последнего нуклеотида в цепи, отрезая «лишние» атомы водорода и кислорода, образующие в результате молекулу воды. Получившуюся связь фосфатной группы с дезоксирибозой называют фосфодиэфирной. Она же используется и при построении молекул РНК.
Из главы 28 ты уже знаешь, что создание дочерних цепей на двух разъединенных матричных цепях ДНК происходит одновременно. И возможно ты обратила внимание на то, что изображается этот процесс часто именно так, как на картинке внизу: одна из расплетенных цепей ДНК вытянута в линию, а другая изогнута петлей.
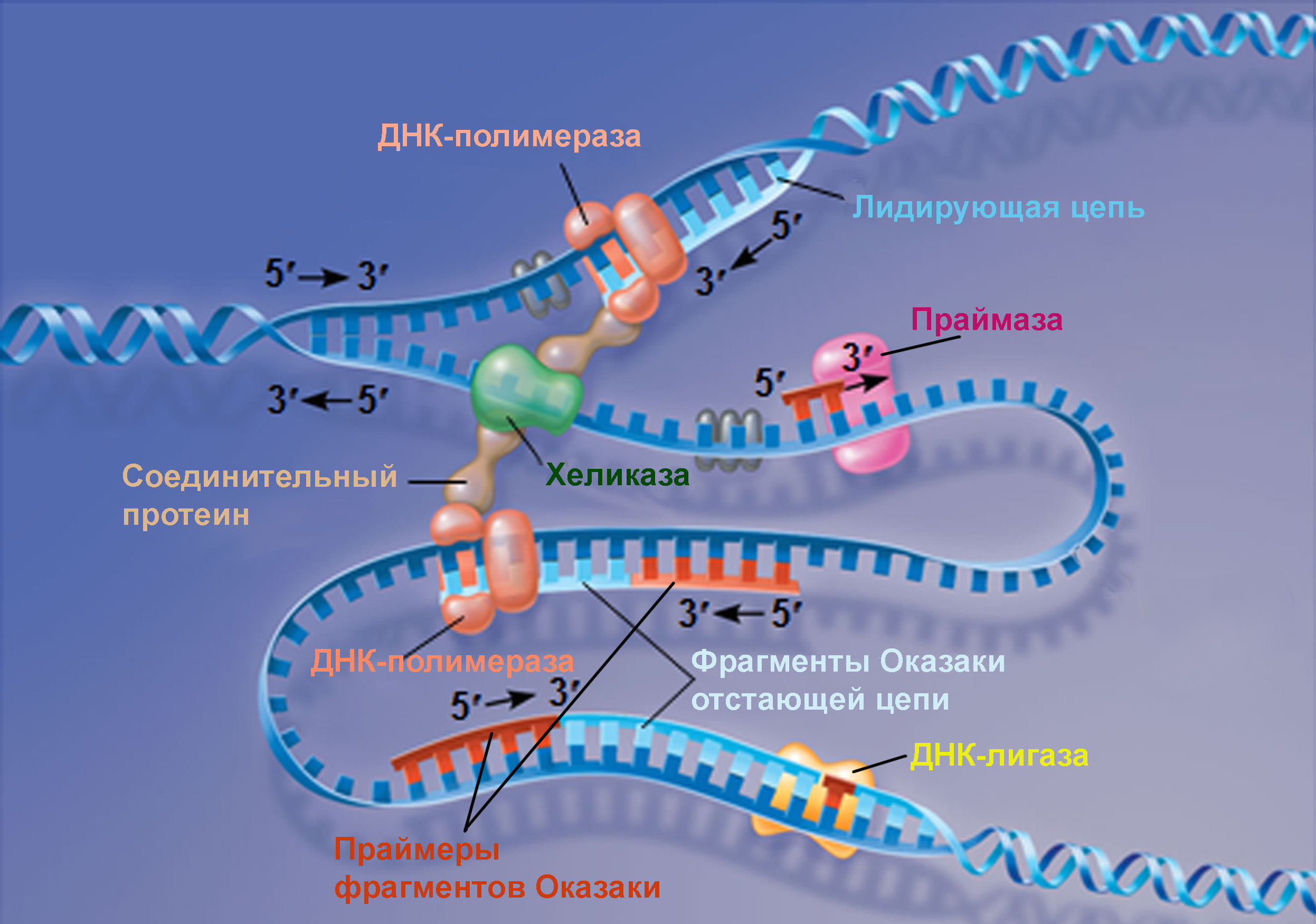
С первой цепью все понятно: вслед за расплетающей материнскую молекулу хеликазой на одну из матричных цепей садится ДНК-полимераза и строит ее, пока не наткнется на идущую навстречу репликативную вилку или на участок терминации, останавливающий копирование. Такую цепь называют лидирующей. К второй, образующей петлю, цепи присоединено больше органелл реплисомы, и причина этого заключается в разнице между двумя цепями ДНК, образующими материнскую молекулу. Две цепи ДНК могут соединиться лишь строго определенным образом: 3′-конец одной соединяется с 5′-концом другой, и если вытянуть двойную спираль в линию, одна из цепей будет лежать в направлении от 5′-конца к 3′-концу, а комплементарная ей будет направлена в противоположную сторону. Такое свойство цепей называется антипараллельностью. На картинке вверху показаны направления всех четырех цепочек ДНК, и матричных, и строящихся.
По каждой из матричных цепей ДНК-полимераза движется от 3′-конца (в этом случае его уместней было бы называть 3′-«начало») к 5′-концу, достраивая антипараллельную дочернюю цепь, которая начинается с 5′-конца и пополняется присоединяемыми с 3′-конца нуклеотидами. Отсюда понятно, что только по одной из материнских цепей ДНК-полимераза может проехать от ее 3′-«начала» до 5′-конца, присоединяя нуклеотиды как только хеликаза отсоединяет от них вторую цепь-матрицу.
Со второй цепью все получается сложнее: ее нельзя взять за начало и строить до конца, потому что вместо 3′-«начала» там 5′-конец. Направление репликации по этой цепи противоположно направлению ее расплетания. Именно поэтому часть этой отстающей цепи начинает строиться только тогда, когда уже значительный участок матричной цепи «оголен»:
=) праймаза создает затравку возле хеликазы, но движется не вслед за ней, а в обратном направлении. На схеме вверху праймаза нарисована как раз в этом месте: она уже установила первый нуклеотид затравки и присоединила к нему второй с его 3′-конца.
=) на затравку, или праймер, садится ДНК-полимераза и достраивает дочернюю цепь, пока не наткнется на готовую двойную спираль. Чаще всего эта спираль является результатом предыдущего прохождения по отстающей цепи праймазы и ДНК-полимеразы, построивших предыдущий участок дочерней цепи. На схеме вверху видно, как эта ДНК-полимераза надстраивает 3′-конец фрагмента отстающей цепи, который скоро дойдет до праймера предыдущего фрагмента. Отстающая цепь строится из множества таких участков «праймер+ДНК», которые называются фрагментами Оказаки. У бактерий фрагменты Оказаки имеют длину около тысячи нуклеотидов, а у эукариот — всего лишь около ста.
=) доходя до праймера, образованного из рибонуклеотидов, ДНК-полимераза млекопитающих вытесняет его с материнской цепи, достраивая полноценную ДНК. Встретив дезоксирибонуклеотид, она отсоединяется от ДНК и перескакивает на более близкий к хеликазе праймер, заполняя нуклеотидами промежуток между ним и только что построенным участком дочерней цепи. У прокариот строящая фрагменты Оказаки ДНК-полимераза не способна вытеснить затравку и покидает ДНК, оставив праймер на месте. С ним работает вспомогательная ДНК-полимераза I прокариот, умеющая вырезать рибонуклеотиды, одновременно заменяя их на дезоксирибонуклеотиды.
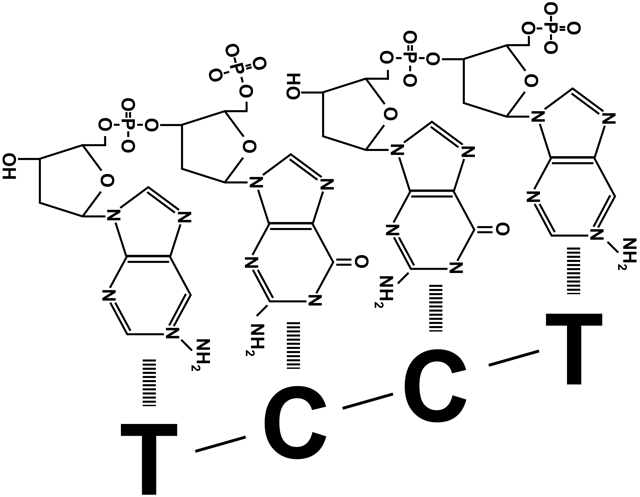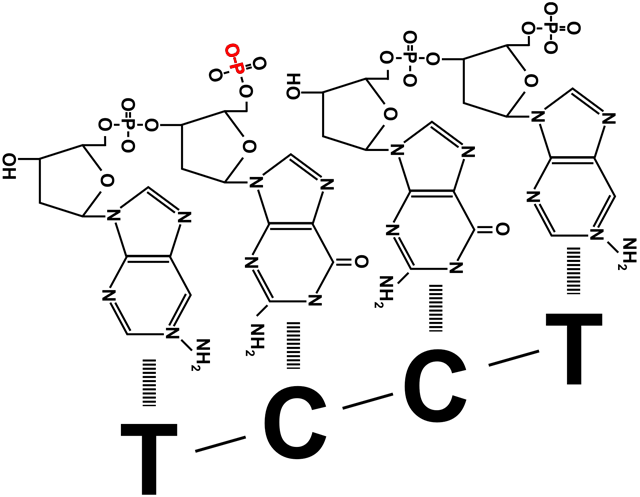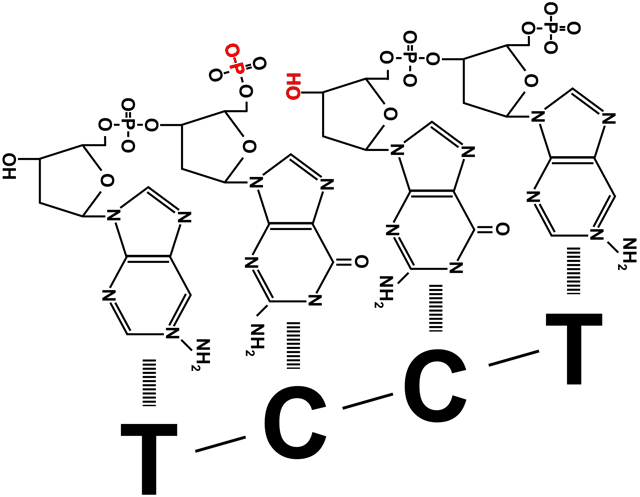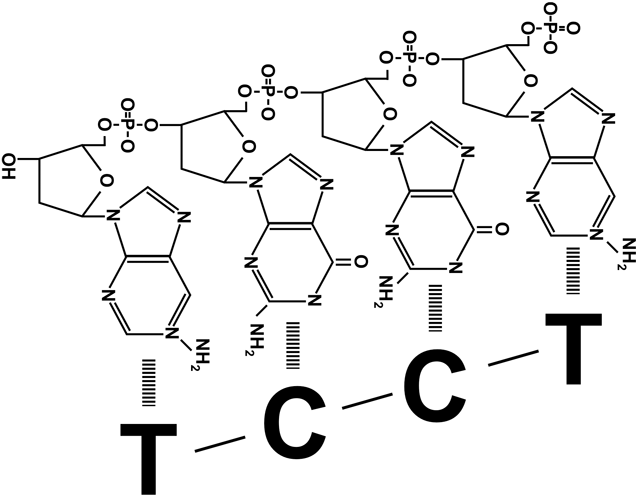
=) вытесненные затравки быстро срезаются с ДНК млекопитающих другими ферментами, после чего остается щель между концом одного фрагмента Оказаки и началом другого. Щель представляет собой не отсутствие одного-двух нуклеотидов, а отсутствие соединения друг с другом фосфата и дезоксирибозы соседних участников цепи. Эту щель «заклеивает» ДНК-лигаза, роль которой не ограничивается этим ее участием в процессе репликации. Цепи ДНК периодически повреждаются, в том числе образуются такие щели между нуклеотидами, или ники. Поэтому ДНК-лигазы часто «инспектируют» ДНК клетки, исправляя повреждения. На схеме справа видно, что они запускают такую же химическую реакцию, что и ДНК-полимеразы при наращивании дочерней цепи. При синтезе ДНК прокариот после замены праймера на дезоксирибонуклеотиды образуется такая же щель между фрагментами Оказаки. Ее устраняет прокариотическая ДНК-лигаза.
Таким образом, несмотря на построение лидирующей и отстающей цепей в противоположных направлениях, они строятся одновременно. И получившаяся в результате соединения ДНК-лигазой многочисленных фрагментов Оказаки, созданных многократно садящимися на материнскую цепь ферментами отстающая цепь ничем не отличается от лидирующей цепи, создание которой стало результатом более простого процесса.
После прочтения этой и предыдущей главы генетической линии ты уже знакома с некоторыми органеллами реплисомы:
=) хеликаза, расплетающая матричную цепь
=) SSB-протеины, удерживающие расплетенные цепи в готовом к репликации состоянии
=) праймазы, начинающие строить дочернюю цепь
=) ДНК-полимеразы, выполняющие основную работу по созданию этой цепи
=) ДНК-лигаза, соединяющая отдельные участки построенной цепи.
Эти органеллы можно назвать основными частями реплисомы. Функции других органелл не менее важны для успешной репликации:
=) в реплисому входит фермент топоизомераза, способный изменять степень закрученности спирали ДНК. Разные виды топоизомераз необходимы и вне процесса репликации для поддержания нитей ДНК в стабильном состоянии: если спираль излишне или недостаточно закручена, это легко может привести к ее повреждению. Один из видов топоизомераз является частью реплисомы: он специализируется на поддержании нормальной степени закрученности того участка ДНК, который предшествует расплетаемому хеликазой.

Чтобы представить, что происходит при расплетании двух образующих спираль нитей ДНК, можно взять веревку, скрученную из двух волокон (такие мы используем для подвязывания веток лимонных туй, нависающих над тропинкой), и начать ее расплетать, взяв по одному волокну в каждую руку и растянув их в стороны. Почти сразу же состоящая из двух волокон часть веревки начнет крутиться, пытаясь компенсировать таким образом расплетание, и даже в таком случае в точке разъединения возникает «перекрученность», мешающая дальнейшему расплетанию волокон. А если веревка длинная, то перекрученность быстро достигнет такой степени, что веревка вообще перестанет раскручиваться. Подергав за концы окончательно запутавшейся веревки, ты легко оценишь важность работы по предотвращению перекручивания нитей ДНК при репликации. Топоизомераза эукариот, выполняющая эту функцию, называется ДНК-гиразой.
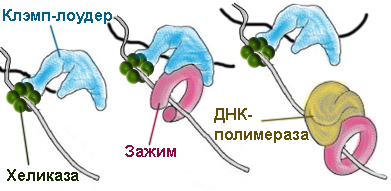
=) на каждой матричной цепи ДНК находится зажим, способный скользить вдоль этой цепи, к которому прикрепляется ДНК-полимераза. Всего в состав реплисомы входят два зажима, по одному для каждой строящейся цепи.
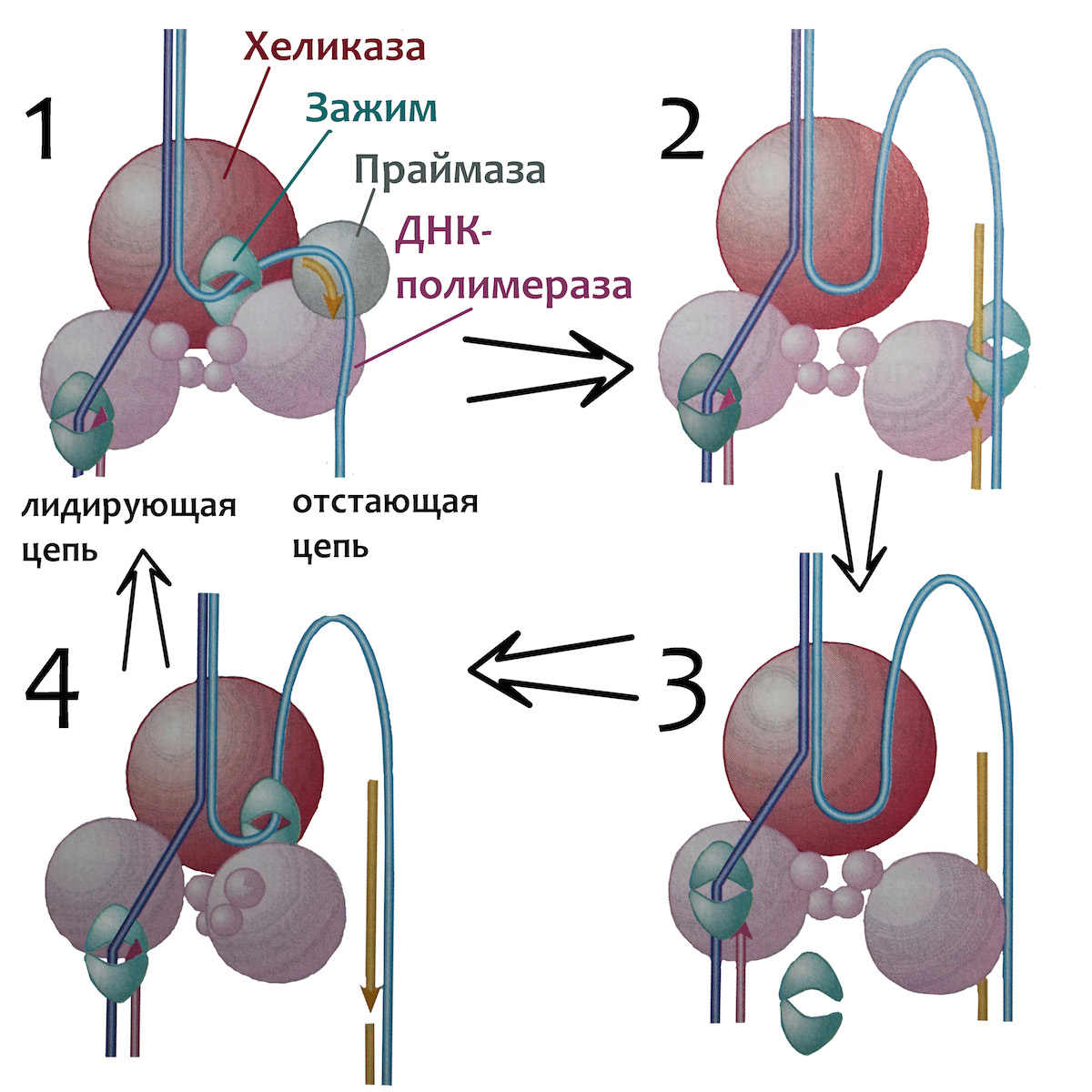
=) Оба зажима раскрывает, накладывает на цепь и защелкивает один клэмп-лоудер, прикрепленный к расплетающей нити матричной ДНК хеликазе. На лидирующую цепь зажим накладывается только один раз, а вот построение отстающей цепи требует постоянного участия клэмп-лоудера: он присоединяет зажим в начале каждого фрагмента Оказаки (рисунок 1), снимает его когда фрагмент достроен (рисунок 3), и переносит к началу следующего фрагмента (рисунок 4).
Итак, подведем итог обеих описывающих воспроизводство ДНК глав учебника:
1. Все ДНК-полимеразы способны создавать цепь ДНК только в одном направлении, присоединяя фосфатную группу нового нуклеотида к 3′-концу строящейся цепи. Так как комплементарные цепи, составляющие молекулу ДНК, противоположно направлены (5′-конец одной соединен с 3′-концом другой цепи), то и синтез дочерних цепей по ним идет в противоположных направлениях. При этом на расплетенном участке по одной (лидирующей) цепи синтез идет непрерывно, а на другой (запаздывающей) создаются отдельные фрагменты Оказаки, строящиеся в направлении, обратном направлению расплетания матричных цепей, то есть «задом наперед». Растущим у фрагментов Оказаки оказывается 3′-конец, как и у лидирующей цепи.
2. Новосинтезированные фрагменты цепи ДНК всегда содержат на 5′-конце несколько рибонуклеотидов. Эту РНК-затравку создает специальный фермент праймаза, который может быть отдельной органеллой реплисомы (у прокариот) или быть субъединицей вспомогательной ДНК-полимеразы (у животных). Предполагают, что использование праймазы объясняется происхождением аппарата репликации ДНК от РНК-полимераз, способных синтезировать исключительно РНК. Также существует функциональное объяснение существования праймеров: в момент инициации синтеза больше вероятность ошибок, поэтому первые установленные в цепь нуклеотиды желательно отличать от остальной новосинтезированной цепи. Использование рибонуклеотидов позволяет безошибочно распознавать затравки, удаляя их впоследствии и накладывая заплатку из дезоксирибонуклеотидов на образовавшийся разрыв в цепи ДНК.
3. В репликации участвует множество органелл, и 3D-анимация дает возможность увидеть их в действии: