



















Всю прошлую главу клетка готовилась к делению, проходя предыдущие фазы клеточного цикла, копируя все, что должно быть скопировано. И наконец-то мы вместе с ней переходим к митозу, или М-фазе — той, в которой она разделится на две одинаковые клетки.
В клетках животных митоз длится один — два часа, а у растений от двух до трех часов. Деление прокариотических (не имеющих ядра) клеток намного быстрее: например, кишечные палочки в благоприятных условиях делятся каждые двадцать минут, но их процесс деления отличается от митоза. Митоз происходит именно в эукариотических клетках, то есть тех, в которых есть ядро. Так как это очень активный процесс, во время которого происходит множество событий, его делят на целых пять фаз: профаза, прометафаза, метафаза, анафаза и телофаза. Звучит как заклинание на каком-то клингонском языке:). У большинства людей, прошедших школьный и университетский концлагеря, это заклинание вызывает тоску и уверенность, что его нужно непременно запомнить, но это ооочень сложно. Так вот сразу1 запоминать его не то что не нужно, а даже вредно — потому что таких похожие слова при попытке запомнить их одновременно только безнадежно перепутаются. Не делай неудобно своему мозгу и просто читай дальше о том, что происходит во время всех этих фаз. Когда каждое из названий будет связано у тебя в памяти с определенными процессами, они перестанут быть такими похожими, и какие-то могут запомниться сами собой.
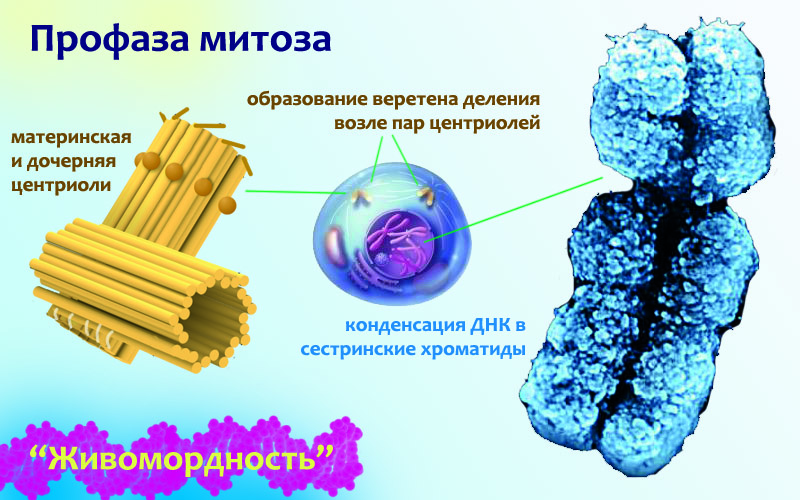
Начинается митоз с профазы, в которой происходят такие процессы:
1. ДНК конденсируется в хромосомы, и теперь их можно увидеть в световой микроскоп. Ты уже знаешь, что каждая из содержащихся в делящейся клетке молекул ДНК была скопирована. Так вот получившиеся две идентичные 2-хспиральные молекулы не расплываются куда попало после копирования, а остаются рядом, скрепленными друг с другом по всей длине специальными протеинами: когезинами. Такую парочку ожидающих деления клетки хромосом называют сестринские хроматиды. Так плотно они сцеплены только в начале профазы, а ближе к концу хроматиды соединяются друг с другом только в одной точке, потому что все когезины отцепились. Только тогда они и образуют похожую на букву Х структуру, которую обычно рисуют не подкованные в генетике художники, если их просят изобразить ДНК (получается, что на рисунке в виде буквы Х обозначено целых 4 ДНК — две двухспиральных молекулы). После отсоединения когезинов единственное, что удерживает сестринские хроматиды вместе, это кинетохоры — другие протеиновые комплексы. Кинетохоры могут присоединяться к ДНК только в особой области, называемой центромерой. На картинке вверху справа — электронная фотография сестринских хроматид, все еще соединенных когезинами. То тонкое место, где они особенно плотно друг к другу прижаты, и есть центромера.

2. Уже понятно, что клетка серьезно настроена разделиться, и все ее силы будут направлены именно на это. А всякие производства молекул будут приостановлены до минимума. Поэтому две крупные клеточные мембранные органеллы, которые используются как раз для этих целей, разбираются на части и становятся фактически нефункциональными. Это эндоплазматический ретикулум (ЭПР) и аппарат Гольджи — заметные клеточные структуры. На картинке слева видно, как они выглядят в неделящейся клетке. А в делящейся клетке во время профазы они превращаются в набор пузырьков, хаотически рассеянных по пространству клетки. После деления, в каждой дочерней клетке эти пузырьки сольются, заново образовав разобранные органеллы.
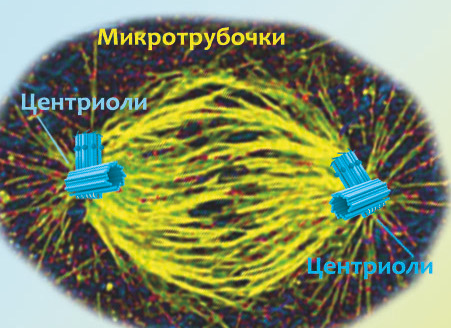
3. Так как клетка будет делиться на две части, необходимо ее самое ценное содержимое — хромосомы — растаскивать по двум противоположным углам. Ну, по условным углам конечно, так как делящиеся клетки чаще всего округлые. И чтобы их растащить, строится удивительная штука: веретено деления. На схеме справа ты видишь его уже готовым: на полюсах клетки находятся пары голубых центриолей, а от них тянутся желтые микротрубочки (в реальных клетках они конечно же не такие яркие, как на схемах). Давай разбираться, что это за штуки, как они устроены и зачем нужны клетке.
Центриоли почему-то есть только у животных, растения и грибы отлично обходятся без них. Изначально в животной клетке находилось две центриоли, но перед делением к каждой из них была построена дочерняя. Получились такие пары цилиндров, перпендикулярно скрепленных друг с другом. Вокруг центриолей находится густой бульон из молекул, которые участвуют в образовании микротрубочек и других протеинов, необходимых для построения веретена деления. Эти области молекулярного бульона, в которых торчат центриоли и из которых вытягиваются микротрубочки, называют центросомами. У растений и грибов, несмотря на отсутствие центриолей, тоже образуются центросомы на полюсах деления, и по размеру они больше, чем те, которые есть в животных клетках. Микротрубочки просто торчат из разных мест этих больших растительно-грибных центросом, в то время как у животных их концы сходятся к центриолям и образуют, таким образом, более прочную структуру. В остальном различий между митозами у эукариот нет.
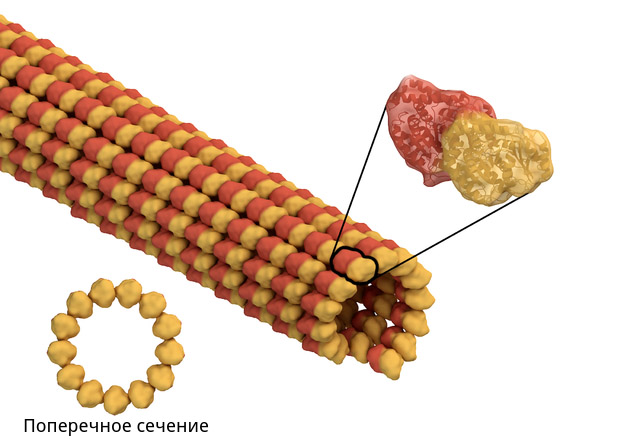
В отличие от центриолей, микротрубочки — это обычная часть клеточного скелета эукариот, они есть в любых обладающих ядрами клетках. Если рассмотреть их поближе (как на картинке слева), то видно, что они собираются из мелких деталей-протеинов. Микротрубочка по английски называется microtubule, отсюда и произошло название составляющих её протеинов: тубулины.
Собираться и разбираться микротрубочки могут с огромной скоростью, подстраивая клеточный скелет под текущие нужды клетки. Например, когда амеба вытягивает свои ложноножки в стремительной погоне за добычей, это как раз результат удлиняющихся внутри ее тела микротрубочек, толкающих вперед клеточную мембрану. И для деления клеток они тоже необходимы, поэтому возле каждой пары центриолей тубулины начинают соединяться в тянущиеся к противоположной паре центриолей микротрубочки. И если хроматиды уже полностью готовы (то есть максимально плотно свернуты), то клетка переходит в следующую фазу митоза — прометафазу.
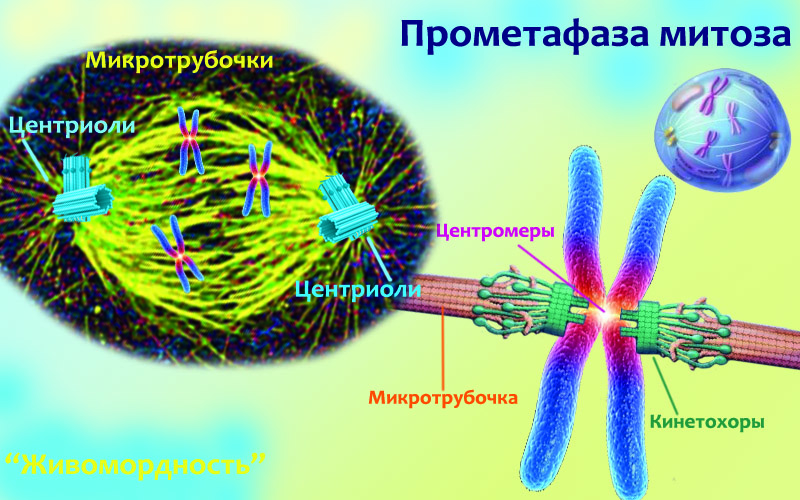
Определить границу между профазой и прометафазой очень легко: в профазе хроматиды все еще сидели в ядре, а в прометафазе оно вдруг исчезло! Правда, будет не совсем верно говорить, что исчезло именно ядро: его содержимое, то есть хроматиды и всякие плавающие возле них молекулы, никуда не делось. Исчезла только удерживающая их в одном месте оболочка. Точно так же, как ЭПР и аппарат Гольджи, оболочка ядра является мембранной органеллой. А значит в нужный момент эта протяженная мембрана распадается на везикулы и создается иллюзия, что она растворилась. Но она все еще находится в клетке, и когда деление закончится, в каждую дочернюю клетку попадет половина тех везикул, на которые распалось ядерная оболочка материнской. Тогда они опять объединятся в единое целое, окружив хромосомы и образовав таким образом ядро дочерней клетки.
Лишенные удерживающей их в ядре оболочки, почувствовавшие свободу хроматиды расплываются по клетке. Если разглядывать этот процесс в микроскоп, то создается впечатление, что они беспорядочно мечутся и прыгают. Но это продолжается недолго: ты еще помнишь, что к центромерам хроматид прикреплены протеины кинетохоры? Вот когда прыгающие хроматиды попадают в область центросомы, строящиеся там микротрубочки цепляются к кинетохорам. На картинке вверху ты видишь ту же пару сестринских хроматид, у которых центромера подсвечена розовым. Только они уже соединены друг с другом не по всей длине, а только в области центромеры (чтобы ее было видно, те части кинетохоров, которые там находятся и обеспечивают это соединение, не нарисованы). И к каждому кинетохору могут присоединиться несколько микротрубочек (на схемах обычно рисуют одну), образуя пучок. Причем присоединенный к хроматидам справа пучок должен соединяться с правой центросомой, и наоборот — левый пучок тянется к левой центросоме. Тогда хроматиды болтаются на длинной нити из микротрубочек, протянутой между двумя полюсами клетки.
Как это получается, и почему оба кинетохора не могут присоединиться к микротрубочкам, идущим от одной и той же центросомы? На самом деле они это могут сделать, и делают, но из-за работы микротрубочек это сразу же опознается как ошибочное присоединение, и один из кинетохоров начинает искать другие микротрубочки, от другой центросомы.
Всякие шевеления микротрубочек при митозе, с помощью которых они выцепляют хроматиды и аккуратно выстраивают их по центру клетки, происходят за счет электромагнинтых взаимодействий между их составными частями — тубулинами. Эти микроскопические штуки роятся, сталкиваясь, мгновенно выстраиваясь в длинную трубочку, которая тут же разрушается, чтобы возникнуть снова, вытянувшись немного в другом направлении. И мне очень сложно удержаться и не начать тут подробно все это описывать — как они взаимодействуют, по каким законам, как именно это беспорядочное столкновение молекул без какого-либо руководства приводит к появлению той упорядоченности, которую мы сейчас увидим в метафазе, как… в общем, я беру себя в руки, а открытый на параграфе про веретено деления учебник из них убираю. И перехожу к следующей фазе митоза: метафазе.
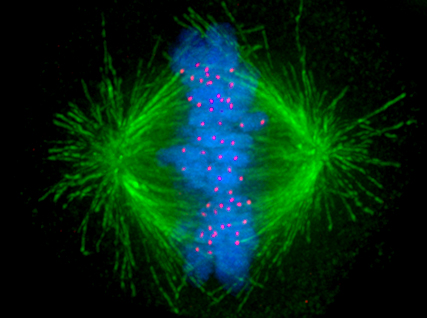
На самом деле про метафазу писать почти нечего: она отличается от прометафазы тем, что хроматиды зафиксированы по центральной линии клетки между полюсами веретена деления, немного колеблясь. Они там висят за счет того, что микротрубочки каждого полюса тянут хроматиды в свою сторону. Пока что кинетохоры прочно удерживают сестринские хроматиды сцепленными, и за счет этого вся система остается в таком вот колеблющемся равновесии. На фотке справа хроматиды подсвечены голубым, кинетохоры красным, а микротрубочки зеленым.
Анафаза — самая короткая из фаз митоза. Сначала кинетохоры, скрепляющие пары сестринских хроматид, вдруг расцепляются — и эти копии молекул ДНК впервые отделяются друг от друга. Самое удивительное, что это происходит одновременно с каждой парой хроматид! Эта синхронность поражает, когда смотришь видео с делящейся клеткой, записанное через микроскоп, возникает уверенность что на самом деле такого быть не может. А оно есть.
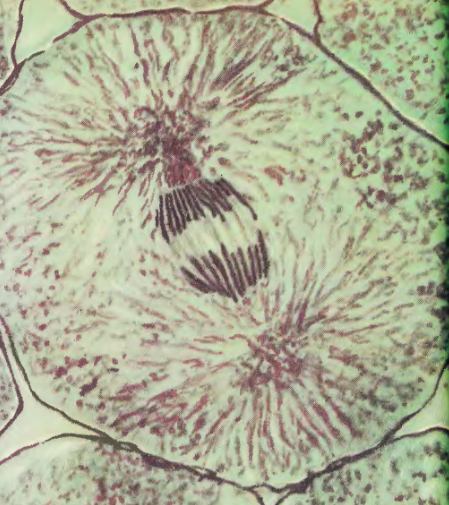
После разрыва связей между хроматидами каждая из пары остается соединенной со своим кинетохором, а через него с пучком микротрубочек. Они и до этого тянули хроматиды к полюсам веретена, но пока хроматиды были соединены в пару, это напоминало безуспешное перетягивание каната. Зато теперь к каждому полюсу деления устремляется свой набор хромосом. Так быстро, что хромосомы приобретают форму буквы V, направленной своим «клювом»-центромерой в сторону полюса.
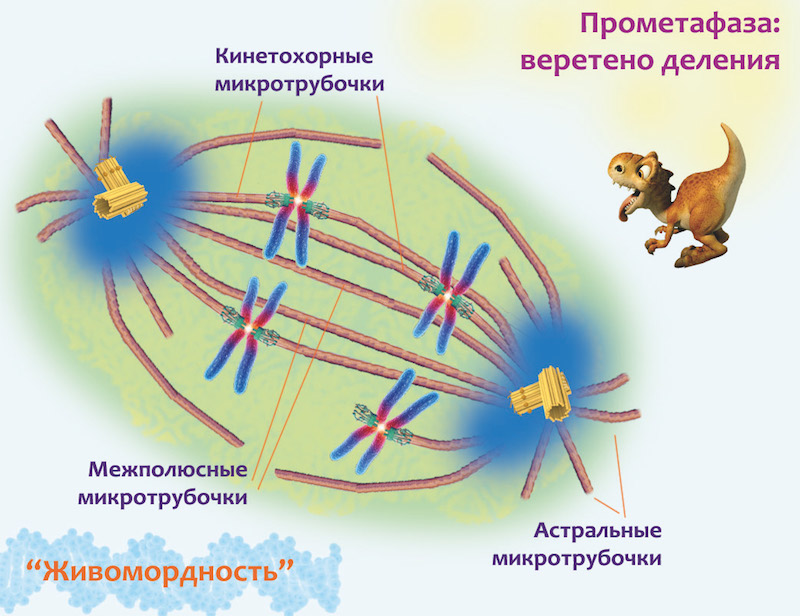
На фотографии слева видно, что хромосомы уже почти приплыли к полюсам веретена, и что эти полюса находятся довольно далеко от клеточных стенок. А для удобства деления стоит их растащить как можно дальше от центра, где будет образовываться перетяжка между образующимися дочерними клетками. И в растаскивании полюсов главную роль играют все те же микротрубочки. Вернись немного назад и посмотри на картинку с веретеном деления в прометафазе: там ты увидишь, что те микротрубочки, которые соединены с хроматидами, называются кинетохорными, и что кроме них есть и другие. Первые — межполюсные — соединяют полюса друг с другом, а вторые протянулись от полюсов деления к клеточной мембране и закрепились за нее. Их называют астральными. Так вот когда хромосомы достигли полюсов деления, кинетохорные трубочки сворачивают свою активность и просто удерживают хромосомы возле полюсов. Зато прохлаждавшиеся до этого межполюсные начинают активно расти, таким образом отпихивая полюса в разные стороны от центра клетки. А астральные микротрубочки наоборот укорачиваются, подтаскивая полюса вместе с болтающимися возле них хромосомами к мембране.
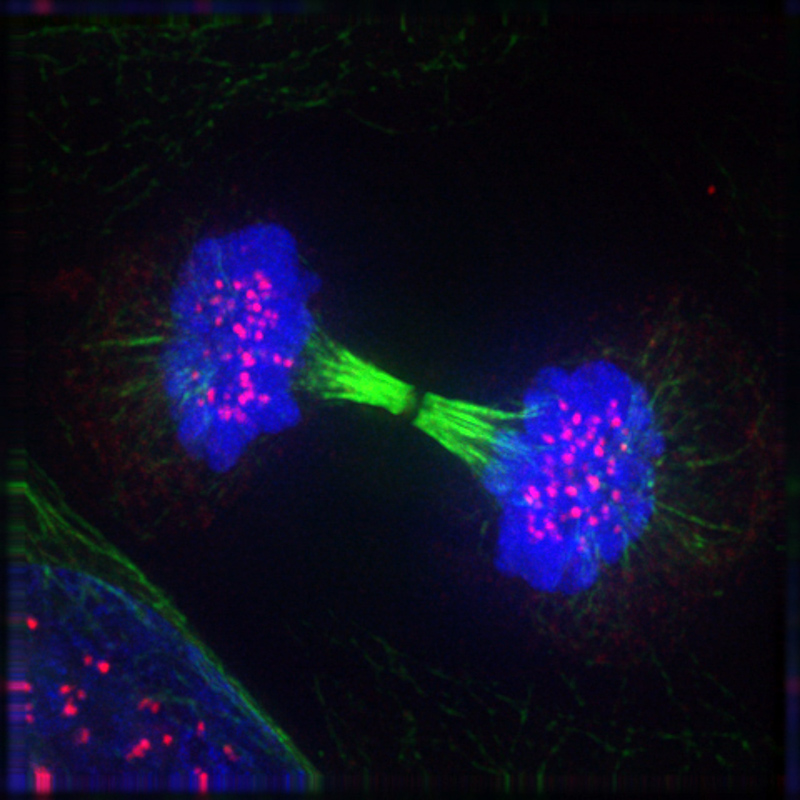
Как только полюса веретена дошли до мембраны и остановились, начинается телофаза. Это последняя фаза митоза, а значит дочерним клеткам уже скоро нужно будет заниматься разными клеточными делами в фазе G1 (если они еще планируют делиться) или в фазе G0 (если они достигли своего клеточного совершенства и приступают к выполнению разных функций для поддержания жизни организма). Подробно про это — в предыдущей главе о подготовке к митозу. Поэтому несмотря на то, что клетка все еще не разделилась, на полюсах веретена начинается активная подготовка к будущему: хромосомы начинают немного распушаться, и к ним прикрепляются те пузырьки, на которые распалась ядерная оболочка материнской клетки. С этого начинается создание ядер дочерних клеток, и оно будет полностью закончено в телофазе. Как только хромосомы полностью окружены ядерной мембраной, они раскручиваются еще сильнее, и в дочерних ядрах образуются ядрышки: более темные из-за обилия роящихся там молекул участки ядра. В ядрышках происходит синтез составных частей для рибосом — очень логично для дочерних клеток начинать свой самостоятельный жизненный цикл именно с этого. Ведь рибосомы необходимы для синтеза протеинов, а без них клетке ну просто никак.
В тот момент, когда у будущих дочерних клеток начинают формироваться ядра, веретено деления разбирается — оно свое дело уже сделало, растащив хроматиды по разным углам клетки. Дальше они уже никуда не денутся, тем более окруженные создаваемыми ядерными оболочками. Фактически в этот момент клетка представляет собой двухъядерного «мутанта», но это не надолго: прежде чем раствориться, веретено деления сыграло еще одну важную роль в процессе митоза. Длинная ось веретена (та, которая соединяет два полюса) используется для определения плоскости, в которой нужно делить клетку пополам. Эта плоскость должна быть перпендикулярна середине длинной оси: она находится там же, где совсем недавно были выстроены в полосу сестринские хроматиды.
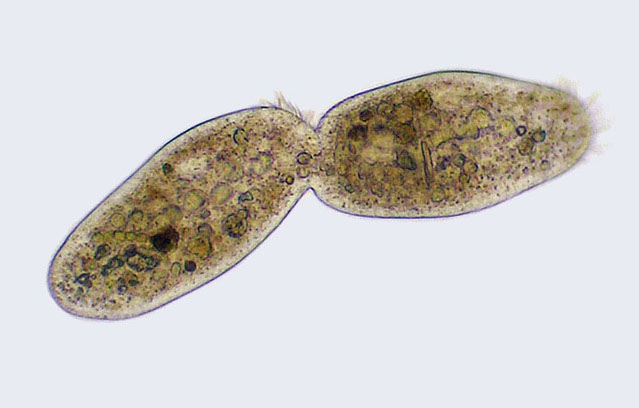
Животные клетки имеют мягкие оболочки, поэтому их деление похоже на перевязывание воздушного шарика веревкой посередине. По диаметру клеточной мембраны с внутренней стороны в месте перетяжки собираются длинные протеиновые нити, образуя кольцо. Они начинают сокращаться, стягивая это кольцо — а за ним тянется и мембрана, потому что нити кольца цепляются за протеины, прочно в ней заякоренные. В конце это больше похоже на затягивание горловины рюкзака, и в какой-то момент узкая перетянутая трубочка клеточной мембраны разрывается посередине.
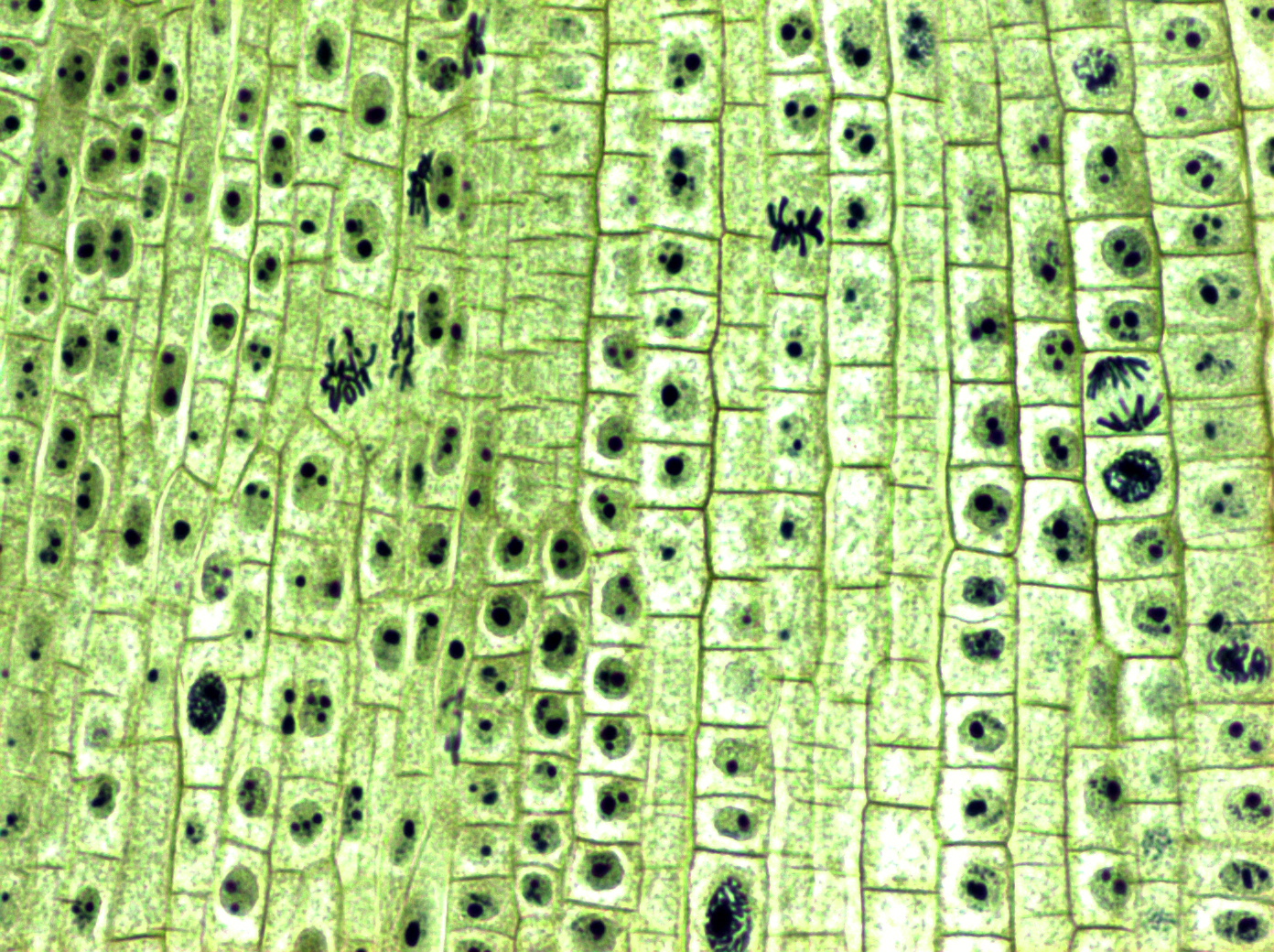
У клеток растений стенки жесткие — как раз за счет этого клетки кожицы лука на фото справа имеют прямоугольную форму. Поэтому при делении между дочерними растительными клетками строится именно такая плотная перегородка.
Итак, митоз закончен. Чтобы вся информация лучше уложилась, еще раз просмотри список названий его стадий и основных происходящих там процессов:
1. Профаза, или Первая фаза:
- ДНК конденсируются, образуются сестринские хроматиды
- приостанавливаются процессы производства протеинов
- разбираются на везикулы мембранные органеллы (ЭПР и аппарат Гольджи)
- строится веретено деления
2. Прометафаза — посередине между профазой и метафазой:
- разбирается на везикулы ядерная оболочка
- освободившиеся из ядра пары сестринских хроматид прыгают по клетке, пока соединяющие их кинетохоры пытаются сцепиться с микротрубочками веретена деления
3. Метафаза:
- хроматиды выстраиваются по центральной плоскости клетки, удерживаемые микротрубочками посередине между полюсами веретена деления
4. Анафаза:
- сестринские хроматиды разделяются и подплывают к полюсам веретена деления
- микротрубочки растаскивают два получившихся набора хромосом по разным сторонам клетки
5. Телофаза — образуются тела новых клеток:
- вокруг каждого набора хромосом строится ядерная оболочка
- начинается производство молекул для постройки рибосом
- дочерние клетки отделяются друг от друга (у животных — перетяжкой мембраны, у растений — построением клеточной стенки) и начинают самостоятельную жизнь.