




















После лирической вводной главы про митохондрию можно наконец рассмотреть ее подробнее, залезая под ее шкурку… собственно, эта шкурка заслуживает первоочередного рассмотрения. Любая клеточная мембрана представляет собой не просто «мешок», в котором болтается содержимое. Это отдельный орган клетки, выполняющий самые разнообразные функции, начиная от закачки в клетку необходимых ей веществ до создания той самой разности потенциалов, которая позволяет археям, бактериям, митохондриям и хлоропластам производить АТФ, запасая таким образом энергию, полученную от расщепления молекул питательных веществ.
У митохондрий есть две мембраны: гладкая внешняя и складчатая внутренняя. Эта складчатость возникла не случайно: чем больше поверхность внутренней мембраны, тем больше на ней разместится энергодобывающих комплексов молекул, которые и будут рассматриваться в этой главе. А чем больше таких комплексов, тем больше энергии может выработать митохондрия.
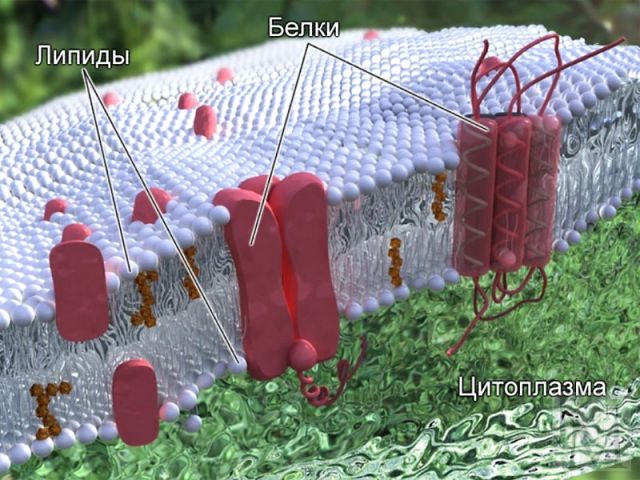
Функция внешней мембраны сравнительно простая: необходимо надежно отгородить митохондрию от цитоплазмы, причем преграда должна быть настолько плотной, чтобы даже одиночный протон без одобрения не просочился. В то же время необходимо постоянно доставлять внутрь митохондрии молекулы строительных веществ, производимых вне ее, и молекулы пищи. И выводить в клетку поставляемую митохондрией АТФ и отходы жизнедеятельности. Для этого в мембрану встроены протеиновые «ворота», причем для каждого типа молекул необходимы специальные, способные пропускать только эти молекулы, устройства.
Таким образом, роль внешней мембраны митохондрии при выработке энергии сводится к функции «забора», и для ее описания достаточно одного абзаца. Но даже поверхностное описание внутренней мембраны займет немало места, ведь именно там происходит все самое интересное, а именно два процесса:
1. манипулирование электронами и протонами для накопления разности потенциалов
2. использование разности потенциалов для сборки молекул АТФ
Эти два процесса тщательно изучаются учеными, и глубина полученной информации позволяет составить очень поверхностное (и за счет этого понятное) описание того, как митохондрия запасает энергию от расщепления питательных веществ, создавая молекулы АТФ:
0. попавшая в митохондрию «еда» (то есть самые разные молекулы, от глюкозы до некоторых жирных кислот) оказывается в матриксе: внутреннем пространстве внутренней мембраны митохондрии. Там она частично окисляется серией химических реакций с участием разнообразных ферментов, передающих измененную до неузнаваемости молекулу по цепи цикла трикарбоновых кислот, или цикла Кребса. Этот цикл является ключевым этапом дыхания всех клеток, использующих кислород. У эукариот его реакции протекают в митохондриях, а у прокариот и архей прямо в цитоплазме. Кроме того участия в накоплении энергии, которое мы сейчас рассматриваем, цикл Кребса поставляет молекулы-предшественники, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения, как аминокислоты, углеводы, жирные кислоты и т.д. Неудивительно, что этот цикл настолько сложен, что на первый взгляд его схема выглядит непознаваемой мешаниной химических процессов. На второй и десятый впрочем тоже.. чтобы не быть голословной, я засунула его схему под этот абзац.
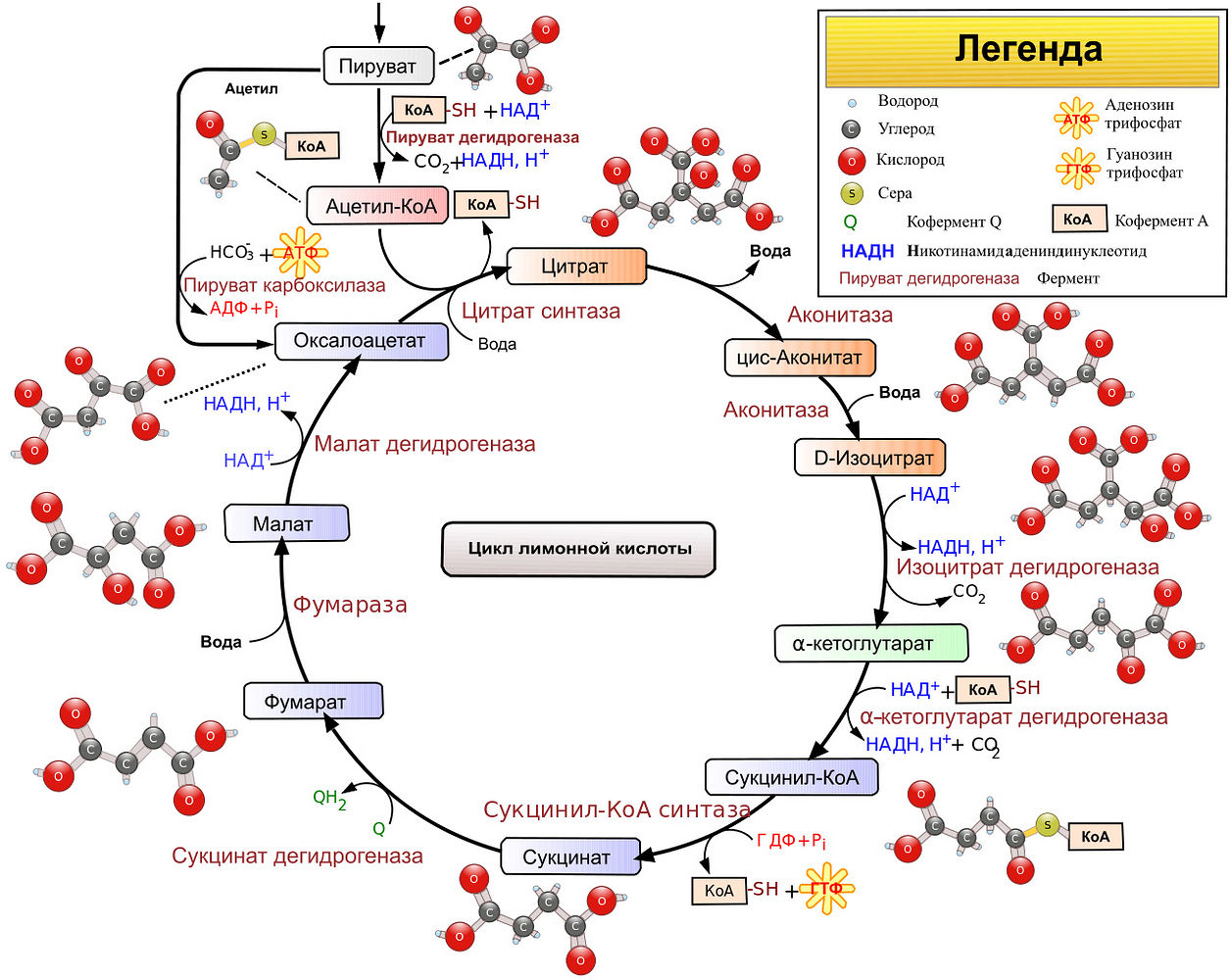
Во время окисления от «еды» отщепляются атомы углерода и кислорода, выводясь наружу в виде углекислого газа, то есть как раз происходит его выдыхание. А в «сухом остатке» цикла Кребса из молекулы например глюкозы образуются две молекулы пирувата и две молекулы АТФ. Казалось бы, АТФ получен — чего еще желать. Но на самом деле это только начало, «пункт ноль» в процессе вырабатывания энергии.
0+. На следующем этапе (назову его 0+, потому что до производства энергии еще далеко) молекулы пирувата и жирных кислот обрабатываются (пируват — одним способом, жирные кислоты — другим), в результате получается одна и та же молекула: ацетил-КоА, в результате окисления которой образуется молекула NADH (сейчас нет необходимости вникать в то, что это за молекулы, поверхностное представление о производстве энергии этого не требует), и вот здесь подготовка для манипуляций с протонами и электронами заканчивается и начинается ее первый этап:
1. NADH легко отдаёт электроны в цепь ферментов, расположенных на кристах (складках внутренней мембраны) митохондрий. Эту электронно-транспортную цепь еще называют «дыхательной цепью» из-за того, что в ней используется кислород в качестве окислителя. Она отщепляет электроны от NADH и в несколько этапов переносит их к кислороду.
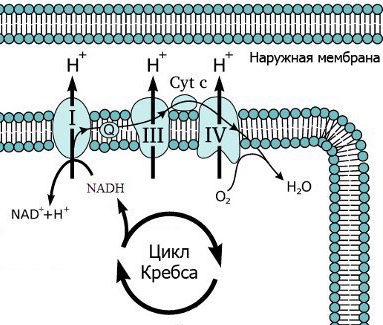
Дыхательная цепь состоит из протеиновых комплексов (на картинке они обозначены как I, III и IV). Первые два содержат атомы металлов, то есть это киборги типа гемоглобина.
Сначала молекула NADH присоединяется к комплексу I, немного сильнее притягивающему электроны содержащихся в ней атомов водорода, чем эта молекула. Этот комплекс отрывает два электрона от молекулы NADH (после этого молекула уплывает), но следующий за ним комплекс в свою очередь немного сильнее притягивает электроны, так что первый недолго наслаждается обладанием и отдает их другому, обозначенному номером III. А в активном центре последнего белкового комплекса IV находится кислород, который гораздо сильнее притягивает электроны, чем оба предыдущих, поэтому они достаются ему. Там из притянувшего электроны кислорода и протонов в итоге образуется вода.
Все эти реакции конечно же не происходят ради создания воды, она здесь образуется как побочный продукт. При каждой передаче электронов от одного белкового комплекса к другому происходит перемещение протонов, которые по специальному транспортному каналу тут же уходят из матрикса через внутреннюю мембрану митохондрии и остаются в межмембранном пространстве. Сначала в комплексе I от молекулы NADH не только отрываются два электрона, которые будут передаваться по дыхательной цепи: оставшиеся без электронов «голые» ядра водорода, то есть протоны, так же отрываются от NADH и используются для активации убихинона (молекулы, переносящей электроны от комплекса I к комплексу III). Предполагают, что активация убихинона так влияет на пространственную форму комплекса I, что он становится способным затянуть в себя протоны, находящиеся в матриксе, и прокачать их через себя.
Каждая передача электронов от одного комплекса дыхательной цепи к другому дает энергию, которая тратится на перенос протонов в межмембранное пространство. Считается, что в процессе транспорта по дыхательной цепи пары электронов через мембрану перекачивается от трёх до шести протонов. В других местах внутренняя мембрана непроницаема для протонов, и внешняя мембрана является мощным «забором», не пропускающим их.
За счет того, что электроны остаются в матриксе, а протоны находятся между мембранами, на внутренней мембране накапливается электрохимический градиент. Даже в случае незаряженных частиц такое неравномерное распределение их в пространстве приводит к их стремлению исправить несправедливость и распределиться равномерно. Имеющийся у протонов электрический заряд существенно увеличивает это стремление, то есть протонный раствор «давит» на обе мембраны митохондрии, пытаясь ворваться внутрь матрикса или вырваться из митохондрии в цитоплазму. И внутрь протоны все-таки пропускают, но не где попало, а там, где это стремление можно использовать (на схеме под этим абзацем это место обозначено цифрой V):
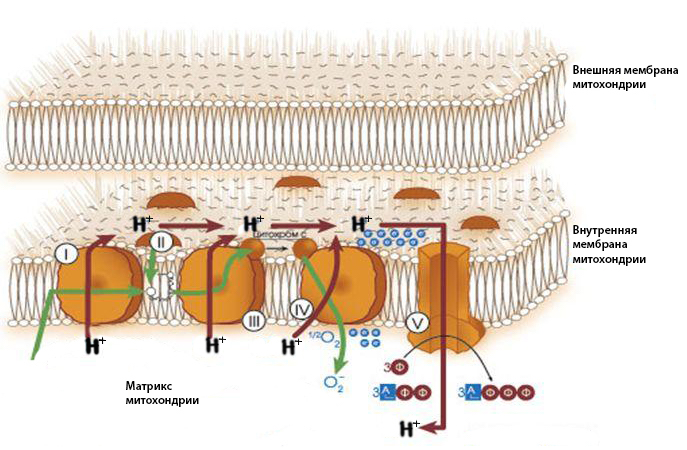
2. Использование протонного градиента происходит на встроенном во внутреннюю мембрану комплексе молекул, который называется АТФ-синтаза. Именно там создаются молекулы АТФ (больше подробностей про запасание энергии в виде АТФ в главе 47).
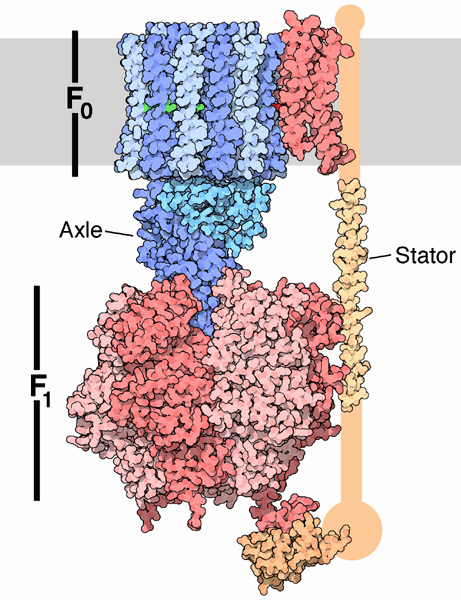

АТФ-синтазы похожи на грибы, которые торчат из внутренней мембраны шляпками в матрикс. Если рассмотреть ее подробно, как на схеме справа, то становятся заметны ее части:
Fо — погруженное в мембрану «основание ножки»
F1 — «шляпка»
Между ними «ножка», обозначенная на картинке как Axle (а почему она называется именно «валом», поймешь позже). На картинке этого не видно, но она проходит и внутри основания ножки, и внутри шляпки. Ее обычно называют γ-субъединицей.
И сбоку какая-то веточка из лесного мусора (stator), прилипшая и к шляпке, и к основанию ножки.
Накопившиеся в межмембранном пространстве протоны пытаются пролезть в матрикс, протискиваясь через основание ножки мимо самой ножки. Эти попытки приводят к тому, что ножка поворачивается — поэтому ее и называют «валом». Шляпка при этом не вращается из-за того, что молекула-stator удерживает ее неподвижной. Основание ножки тоже не двигается. На большинстве найденных в Интернете схем и даже 3D-видео движение этих частей изображается неправильно, возможно для большей наглядности (ведь вращение закрытой шляпкой и основанием ножки не так заметно), возможно из-за того, что они уже устарели.
Получается, что энергия, запасенная в виде концентрации протонов в межмембранном пространстве, расходуется на вращение ножки АТФ-синтазы. Что же дает это вращение?

Шляпка F1 состоит из шести «долек»: чередующихся друг с другом α- и β-субъединиц (по 3 каждого вида). Неважно, как именно они устроены, главное то, что они способны присоединять к себе две молекулы: АДФ (отличающийся от АТФ только отсутствием третьей фосфатной группы) и ту самую недостающую фосфатную группу.
Просто поместив эти молекулы рядом, собрать АТФ невозможно: необходимо прижать их друг к другу, затратив на это энергию. И как раз во время вращения γ-субъединицы («ножки») эта энергия и прикладывается. По мере вращения γ-субъединицы в трех местах единицы F1 сначала присоединяются АДФ и фосфатная группа, потом они объединяются в молекулу АТФ, потом готовая молекула «выкидывается» из АТФазы:
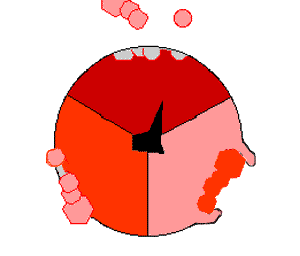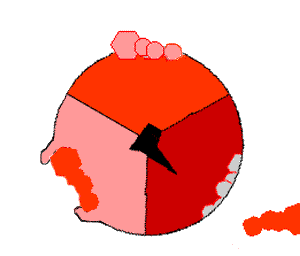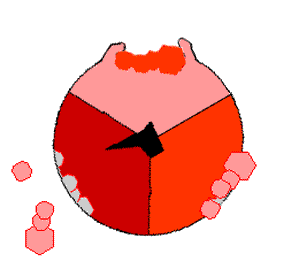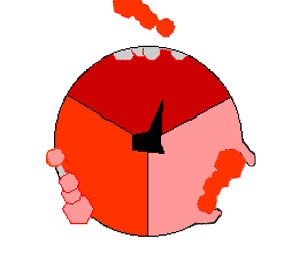
На картинке справа вращающаяся γ-субъединица обозначена черным, АДФ и фосфатная группа — розовым, АТФ — красным.
Все это гораздо больше похоже на описание некоего мотора, механического устройства на протонной тяге, чем на неотъемлемую часть живых существ, от бактерий и архей до человека. Но именно такими «моторами» усеяны внутренние мембраны каждой из живущих в тебе митохондрий.

Собираемые здесь молекулы АТФ используются для любого твоего движения, физического или психического. Без симбионтов-митохондрий возможности клетки жестко ограничены: не будет ни увеличения размера, как у эукариотических гигантов-одноклеточных, ни многоклеточности, ни появления сознания в том виде, в котором мы его у себя наблюдаем. Без митохондрий предельный уровень сложности организма остается на уровне бактерий и архей. За миллионы лет эволюции они непрерывно изменялись, приспосабливаясь к новым условиям, но чего-то большего на их базе не возникло. Сравни это с разнообразием эукариот, которое стало возможным благодаря вот этому: